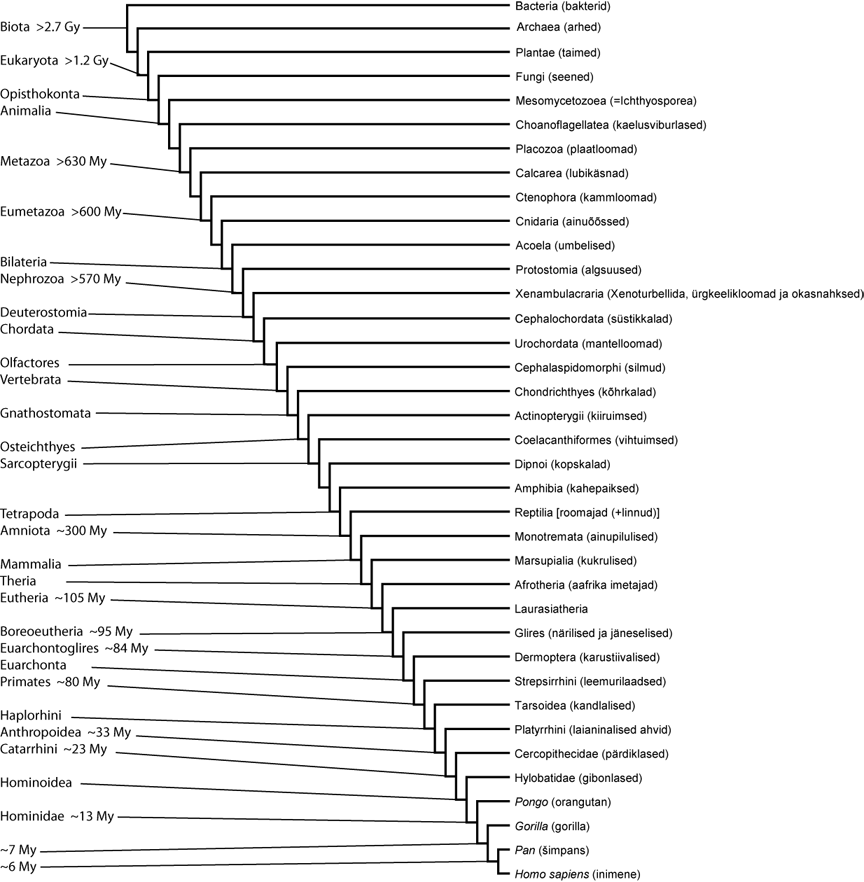
Joonis 4. Inimese positsioon eluslooduse fülogeneesi puus ("Elupuus"). Kladogramm Lang et al. (2002), Steenkamp et al. (2006), Dellaporta et al. (2006), Wallberg et al. (2004), Baguñà & Riutort (2004), Bourlat et al. (2006), Cameron et al. (2000), Meyer & Zardoya (2003), Murphy et al. (2001), Ross (2000) järgi. Erinevate rühmade lahknemise ajad on Lepland & Lepland (2002), Porter (2004), Peterson & Butterfield (2005), Springer et al. (2003), Glazko & Nei (2003) järgi. Kommentaare vt. allpool. Gy - miljard aastat tagasi; My - miljon aastat tagasi.
Ülaltoodud kladogrammi abil on tahetud näidata inimese positsiooni selles võimalikult täpselt ja samas tuues välja võimalikult vähe harusid, milletõttu ongi see nii ebasümmeetriline (kuna ühegi sõsarrühma siseseid fülogeneetilisi suhteid pole näidatud). Inimese sõsarrühmaks (lähimaks sugulaseks) on perekond Pan (Pan troglodytes+Pan paniscus), neile (Homo+Pan) omakorda gorilla jne. Siiski on teadmiste vähesuse tõttu välja jäetud mitmed taksonid, kes võiksid potentsiaalselt pisut seda kladogrammi täiendada/muuta. Taksonid, kes said välja jäetud, on järgmised: tupaialised (Scandentia), napihambulised (Xenarthra), pihklased (Myxini), Nemertodermatida, päriskäsnad (Demospongiae), klaaskäsnad (Hexactinellida), Homoscleromorpha (klassifitseeriti varem päriskäsnade klassi) ja mitmed protistide rühmad (Amoebozoa, Rhizaria, Chromalveolata, Excavata ja mõned ainuraksed tagaviburlased).
On selgusetu, kes on Boreoeutheria sõsarrühmaks, kas Xenarthra, Afrotheria või Xenarthra+Afrotheria klaad, seetõttu jätsin meelevaldselt välja napihambulised. Pihklaste ja silmude fülogeneetiline positsioon selgroogsete seas on vastuoluline. Morfoloogiliste andmete kohaselt on silmud lõugsuuste (Gnathostomata) sõsarrühmaks (seetõttu eelistasin kladogrammil silme pihklastele) (vt. Meyer & Zardoya, 2003), kuid mitmed molekulaarsed analüüsid kipuvad toetama traditsioonilist sõõrsuude (Cyclostomata) klaadi (Delarbre et al., 2002; Takezaki et al., 2003; Delsuc et al., 2006), milletõttu peaks ülal toodud kladogrammil silmude haru asendama sõõrsuude omaga. Kuna Acoelomorpha (Acoela+Nemertodermatida) monofüleetilisus pole kinnitust leidnud molekulaarsete andmete põhjal (e. g. Wallberg et al., 2004; Glenner et al., 2004), siis on kladogrammil toodud ainult umbelised (Acoela). Käsnade hõimkond on suure tõenäosusega parafüleetiline, kuid kahjuks on veel jäänud ebaselgeks, milline käsnade rühm võiks olla sõsarrühmaks pärishulkraksetele (Eumetazoa) (vt. näit. Kruse et al., 1998; Borchiellini et al., 2001; Peterson & Eernisse, 2001; Borchiellini et al., 2004) ja seetõttu on joonisel toodud ainult lubikäsnad (Calcarea). Kuna ainuviburiliste ja kaheviburiliste (vt. eukarüootide fülogeneesi) monofüleetilisus pole kindel, siis on eukarüootidest lisaks tagaviburlastele toodud ainult taimed (Plantae). Päristuumsete sõsarrühmaks on pandud arhed (Archaea) ja arhede/eukarüootide klaadile omakorda bakterid (Bacteria) (e. g. Woese et al., 1990).
Xenoturbella bocki paigutati 18S rDNA (SSU) ja kahe mitokondri geeni põhjal teissuuste hulka 2003. aastal (Bourlat et al., 2003). SSU (ribosoomi väikse alaühiku RNA) geeni analüüs näitas, et Xenoturbella võiks olla Ambulacraria (ürgkeelikloomad ja okasnahksed) sõsarrühmaks. Sellele leiti kinnitus Bourlat et al. (2006) poolt, kellel oli kasutada juba terve mitokondri genoom ja 63 rakutuuma valgugeeni. Nad panid Xenoturbellida ja Ambulacraria klaadile nimeks Xenambulacraria.
Süstikkalad (Cephalochordata) ei ole molekulaarsete andmete põhjal selgroogsete sõsarrühmaks ja nende kuulumine keelikloomade hulka (nagu näidatud joonisel) ei olegi väga kindel. Nimelt kasutasid Delsuc et al. (2006) fülogeneetilistes analüüsides 146 tuumageeni järjestusi ja demonstreerisid veenvalt, et selgroogsete sõsarrühmaks on tegelikult mantelloomad (Urochordata) ja mitte süstikkalad. Süstikkalad võivad Delsuc et al. (2006) järgi olla hoopis lähemas suguluses okasnahksetega kui teiste keelikloomadega. Aga kuna analüüsides oli ainult üks okasnahkne, üks süstikkala ja ürgkeelikloomad puudusid üldse, siis ei pidanud Delsuc et al. (2006) seda tulemust loomulikult väga tõsiseltvõetavaks. Ja tõepoolest, kui analüüsidesse lisati üks ürgkeelikloom (Hemichordata; okasnahksete sõsarrühm), siis oli süstikkalade paiknemine fülogeneesis okasnahksete lähedal oluliselt vähem toetatud (Marlétaz et al., 2006). Samas on keelikloomade monofüleetilisus kahtluse alla seatud ka embrüoloogilistele tunnustele tuginedes (Raineri, 2006). Bourlat et al. (2006), kes aga lisasid Delsuc et al. (2006) andmestikule lisaks Xenoturbella (63 geeni), ürgkeeliklooma (153 geeni) ja veel ühe okasnahkse (67 geeni) järjestused, leidsid et keelikloomad võiksid siiski olla monofüleetilist päritolu (statistiline toetus sellele hüpoteesile oli aga nõrk).
Süstikkalasid on peaaegu universaalselt peetud selgroogsete sõsarrühmaks, kuna nad meenutavad kalasid ja neil on selgroogsetega mitmeid ühiseid tunnuseid: külgmised lihasväädid, mis on ristvaheseintega (müoseptidega) jaotunud lihassegmentideks (müomeerideks); üldehituselt sarnane vereringe süsteem ning segmenteerunud organite innervatsioon selgmiste, seljaajust algavate närvide poolt. Nüüd on siis välja tulnud, et neid tunnuseid ei saa pidada süstikkalade ja selgroogsete sünapomorfideks vaid need on hoopis sümplesiomorfsed - mantelloomad on need tunnused lihtsalt kaotanud oma spetsialiseerunud eluviisi tõttu. Lisaks molekulaarsetele andmetele (Delsuc et al., 2006; Bourlat et al., 2006) toetavad klaadi Olfactores ka mõned morfoloogilised analüüsid (Ruppert, 2005), võimalike neuraalharja (nn. "neljas looteleht") rakkude olemasolu mantelloomadel (süstikaladel pole neid leitud) (Jeffery et al., 2004), kadheriini molekulide struktuur (need molekulid osalevad rakkude vaheliste adherents-liiduste moodustamisel; Oda et al., 2002) ja 6-8 aminohappe pikkune insertsioon ühes kollageeni geenis (Wada et al., 2006).
Natuke ka joonisel toodud erinevate organismide tekkimise aegade kohta.
Elu alguse dateerimise üle on vaieldud. Viimasel ajal on kahtluse alla seatud tõendid, nagu eksisteerisid juba 3,5 miljardit aastat tagasi tsüanobakterid ning vanimateks kindlalt tõestatud elu jälgedeks peetakse seetõttu hoopis orgaanilisi molekule 2,7 miljardi aasta tagusest ajast (Lepland & Lepland, 2002). Hiljuti tehtud uurimuses tõid Tice & Lowe (2004) tõendeid, et fotosünteesivad organismid võisid siiski eksisteerida ~3,4 miljardit aastat tagasi, kuid ilmselt ei olnud tegemist tsüanobakteritega, vaid mitteoksügeensete fotosünteesivate mikroobidega. Tulemused sobivad hästi Cavalier-Smith (2002) vaadetega, kes arvab, et just sellised bakterid olid kõige esimesed ja tsüanobakterid on tekkinud tunduvalt hiljem (~2 miljardit aastat tagasi).
Küsimusele millal tekkisid eukarüoodid pole ka rahuldavat vastust leitud. Üldiselt peetakse kõige veenvamateks vanimateks eukarüootseteks fossiilideks 1,2 miljardi aasta vanuseid leide (punavetikad) (Simpson & Roger, 2004; Porter, 2004). Kuid Cavalier-Smith (2002) interpreteerib neid kui tsüanobaktereid ja arvab, et eukarüoodid tekkisid ja nende kiire evolutsioon toimus alles ~850 miljonit aastat tagasi. Cavalier-Smith (2002) on vist enam-vähem ainuke, kes pooldab eukarüootide nii hilist teket ning Javaux et al. (2003) on tema interpretatsioonid kahtluse alla seadnud. Javaux et al. (2003) kirjeldavad mitmeid ~1,5 miljardi aasta vanuseid fossiile, kes kõige tõenäolisemalt on eukarüoodid, kuid keda ei ole võimalik kindlalt paigutada ühessegi protistide rühma.
Seoses esikloomaliste erinevate rühmade lahknemiste aegadega tuleks joonisel sisse viia ehk mõned parandused. Steiper & Young (2006) leidsid, kasutades u. 60 000 bp tuuma DNA-d, et primaadid tekkisid ~77 miljonit aastat tagasi (Ma), ahvilised (Anthropoidea) ~43 Ma, kitsaninalised ahvid (Catarrhini) ~30 Ma ja inimahvid (Hominidae) 18 Ma. Molekulaarse kella kalibreerimisel kasutasid Steiper & Young (2006) muuhulgas inimise ja šimpansi lahknemist 6–7 Ma. Selle kalibreeringu aluseks oli Sahelanthropus tchadensis, mida on peetud varaseimaks (pärast šimpansist lahknemist) inimesega ühte evolutsiooni harusse kuuluvaks fossiiliks (Brunet et al., 2002; 2005). Kuid selles võib kahelda, kas Sahelanthropus tchadensis ikka on inimesega lähemas suguluses kui teiste inimahvidega. Pigem on ta lahknenud juba enne šimpansi ja inimese ühist klaadi, võibolla isegi enne Gorilla+Pan+Homo klaadi.
Viited
Baguñà, J. & Riutort, M. 2004. Molecular phylogeny of the Platyhelminthes. Canadian Journal of Zoology 82: 168–193.
Borchiellini, C., Manuel, M., Alivon, E., Boury-Esnault, N., Vacelet, J. & Le Parco, Y. 2001. Sponge paraphyly and the origin of Metazoa. Journal of Evolutionary Biology 14: 171–179.
Borchiellini, C., Chombard, C., Manuel, M., Alivon, E., Vacelet, J. & Boury-Esnault, N. 2004. Molecular phylogeny of Demospongiae: implications for classification and scenarios of character evolution. Molecular Phylogenetics and Evolution 32: 823–837.
Bourlat, S. J., Nielsen, C., Lockyer, A. E., Littlewood, D. T. J. & Telford, M. J. 2003. Xenoturbella is a deuterostome that eats molluscs. Nature 424: 925–928.
Bourlat, S. J., Juliusdottir, T., Lowe, C. J., Freeman, R., Aronowicz, J., Kirschner, M., Lander, E. S., Thorndyke, M., Nakano, H., Kohn, A. B., Heyland, A., Moroz, L. L., Copley, R. R. & Telford, M. J. 2006. Deuterostome phylogeny reveals monophyletic chordates and the new phylum Xenoturbellida. Nature 444: 85–88.
Cameron, C. B, Garey, J. R. & Swalla, B. J. 2000. Evolution of the chordate body plan: New insights from phylogenetic analyses of deuterostome phyla. Proceedings of the National Academy of Sciences. USA 97: 4469-4474. [pdf]
Cavalier-Smith, T. 2002. The neomuran origin of archaebacteria, the negibacterial root of the universal tree and bacterial megaclassification. International Journal of Systematic and Evolutionary Microbiology 52: 7–76. [pdf]
Delarbre, C., Gallut, C., Barriel, V., Janvier, P. & Gachelin, G. 2002. Complete Mitochondrial DNA of the Hagfish, Eptatretus burgeri: The Comparative Analysis of Mitochondrial DNA Sequences Strongly Supports the Cyclostome Monophyly. Molecular Phylogenetics and Evolution 22: 184-192.
Dellaporta, S. L., Anthony Xu, A., Sagasser, S., Jakob, W., Moreno, M. A., Buss, L. W. & Schierwater, B. 2006. Mitochondrial genome of Trichoplax adhaerens supports Placozoa as the basal lower metazoan phylum. Proceedings of the National Academy of Sciences. USA 103: 8751–8756. [pdf]
Delsuc, F., Brinkmann, H., Chourrout, D. & Philippe, H. 2006. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 439: 965–968.
Glazko, G. V. & Nei, M. 2003. Estimation of Divergence Times for Major Lineages of Primate Species. Molecular Biology and Evolution 20: 424-434. [pdf]
Glenner, H., Hansen, A. J., Sørensen, M. V., Ronquist, F., Huelsenbeck, J. P. & Willerslev, E. 2004. Bayesian Inference of the Metazoan Phylogeny: A Combined Molecular and Morphological Approach. Current Biology 14: 1644-1649; Erratum: Current Biology 15: 392-393 (2005).
Javaux, E. J., Knoll, A. H. & Walter, M. 2003. Recognizing and Interpreting the Fossils of Early Eukaryotes. Origins of Life and Evolution of the Biosphere 33: 75-94.
Jeffery, W. R., Strickler, A. G. & Yamamoto, Y. 2004. Migratory neural crest-like cells form body pigmentation in a urochordate embryo. Nature 431: 696-699.
Kruse, M., Leys, S. P., Müller, I. M. & Müller, W. E. G. 1998. Phylogenetic Position of the Hexactinellida Within the Phylum Porifera Based on the Amino Acid Sequence of the Protein Kinase C from Rhabdocalyptus dawsoni. Journal of Molecular Evolution 46: 721-728.
Lang, B. F., O'Kelly, C., Nerad, T., Gray, M. W. & Burger, G. 2002. The Closest Unicellular Relatives of Animals. Current Biology 12: 1773–1778.
Lepland, A. & Lepland, A. 2002. Elu jäljed ürgses kivis. Eesti Loodus 53: 8–14.
Marlétaz, F., Martin, E., Perez, Y., Papillon, D., Caubit, X., Lowe, C. J., Freeman, B., Fasano, L., Dossat, C., Wincker, P., Weissenbach, J. & Le Parco, Y. 2006. Chaetognath phylogenomics: a protostome with deuterostome-like development. Current Biology 16: R577–R578.
Meyer, A. & Zardoya, R. 2003. Recent Advances in the (molecular) phylogeny of Vertebrates. Annual Review of Ecology, Evolution, and Systematics 34: 311-338.
Murphy, W. J., Eizirik, E., O'Brien, S. J., Madsen, O., Scally, M., Douady, C. J., Teeling, E., Ryder, O. A., Stanhope, M. J., de Jong, W. W. & Springer, M. S. 2001. Resolution of the Early Placental Mammal Radiation Using Bayesian Phylogenetics. Science 294: 2348-2351.
Oda, H., Wada, H., Tagawa, K., Akiyama-Oda, Y., Satoh, N., Humphreys, T., Zhang, S. & Tsukita, S. 2002. A novel amphioxus cadherin that localizes to epithelial adherens junctions has an unusual domain organization with implications for chordate phylogeny. Evolution & Development 4: 426–434.
Peterson, K. J. & Butterfield, N. J. 2005. Origin of the Eumetazoa: Testing ecological predictions of molecular clocks against the Proterozoic fossil record. Proceedings of National Academy of Science. USA 102: 9547-9552. [pdf]
Peterson, K. J. & Eernisse, D. J. 2001. Animal phylogeny and the ancestry of bilaterians: inferences from morphology and 18S rDNA gene sequences. Evolution & Development 3: 170-205.
Porter, S. M. 2004. The fossil record of early eukaryotic diversification. Paleontological Society Papers 10: 35-50. [pdf]
Raineri, M. 2006. Are protochordates chordates? Biological Journal of the Linnean Society 87: 261–284.
Ross, C. F. 2000. Into the light: the origin of Anthropoidea. Annual Review of Anthropology 29: 147-194.
Ruppert, E. E. 2005. Key characters uniting hemichordates and chordates: homologies or homoplasies? Canadian Journal of Zoology 83: 8–23.
Simpson, A. G. B. & Roger, A. J. 2004. The real ‘kingdoms’ of eukaryotes. Current Biology 14: R693–R696.
Springer, M. S., Murphy, W. J., Eizirik, E. & O'Brien, S. J. 2003. Placental mammal diversification and the Cretaceous-Tertiary boundary. Proceedings of National Academy of Science. USA 100: 1056-1061. [pdf]
Steenkamp, E. T., Wright, J. & Baldauf, S. L. 2006. The Protistan Origins of Animals and Fungi. Molecular Biology and Evolution 23: 93–106. [pdf]
Steiper, M. E. & Young, N. M. 2006. Primate molecular divergence dates. Molecular Phylogenetics and Evolution 41: 384–394.
Takezaki, N., Figueroa, F., Zaleska-Rutczynska, Z & Klein, J. 2003. Molecular Phylogeny of Early Vertebrates: Monophyly of the Agnathans as Revealed by Sequences of 35 Genes. Molecular Biology and Evolution 20: 287-292. [pdf]
Tice, M. M. & Lowe, D. R. 2004. Photosynthetic microbial mats in the 3,416-Myr-old ocean. Nature 431: 549–552.
Wada, H., Okuyama, M., Satoh, N. & Zhang, S. 2006. Molecular evolution of fibrillar collagen in chordates, with implications for the evolution of vertebrate skeletons and chordate phylogeny. Evolution & Development 8: 370–377.
Wallberg, A., Thollesson, M., Farris, J. S. & Jondelius, U. 2004. The phylogenetic position of the comb jellies (Ctenophora) and the importance of taxonomic sampling. Cladistics 20: 558–578.
Woese, C. R, Kandler, O. & Wheelis, M. L. 1990. Towards a Natural System of Organisms: Proposal for the Domains Archaea, Bacteria, and Eucarya. Proceedings of National Academy of Science. USA 87: 4576-4579. [pdf]