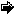
| | | 7.3 Eutrophication and Phytoplankton: the Mass Balance Approach |
Operationally, two different approaches have been applied in analyzing the effects of eutrophication on phytoplankton: a mass balance approach and an organismal approach. The eutrophication classification schemes discussed above, i.e. the increased rate of supply of organic carbon to an ecosystem (Nixon); the ratio of particulate to dissolved organic carbon, the soluble amino acid turnover times (Seki & Iwami); winter dissolved inorganic nitrogen levels and summer chlorophyll (= biomass) levels (CSTT) are mass balance approaches. These classifications emphasize the abundance (yield) of phytoplankton that develops in response to added nutrient (dose). Conceptually, the mass balance approach applies the photosynthesis-respiration reaction (Eq. 1)in combination with the stoichiometric relationships (Redfield Ratio) that occur between nutrients (nitrogen, phosphorus) and the photosynthesis and respiration of organic matter. In this relationship, nitrogen (N) and phosphorus (P) are bound and released during the synthesis and respiration of organic carbon (C) which, in turn, result either in the production or utilization of oxygen (O):
O:C:N:P
The quantities of the four elements processed in this biochemistry, known as the Redfield Ratio, can be expressed in terms of atoms (Eq. 1), or by weight (Eq. 1):
O:C:N:P=212:106:16:1
O:C:N:P=109:41:17.2:1
Thus, during photosynthesis (primary production) for every atom of P assimilated, 16 atoms of N will be assimilated and 106 atoms of C fixed into organic matter (from CO2 -- see Eq. 1), liberating 212 atoms of O. This assimilation of N and P leads to phytoplankton growth (i.e. biomass = carbon, chlorophyll, etc.) and oxygenates the watermass. Ideally, from a knowledge of the amount of N and P available, for example the DAIN concentrations recommended by CSTT, one can estimate the amount of C that would be produced to evaluate potential eutrophication effects. And, from the biomass level, i.e. the summer chlorophyll (= biomass) levels applied by CSTT as an index of eutrophication status, one can estimate the amount of N and P needed to produce and sustain that biomass level. In that case, one needs to know the C:Chlorophyll a ratio, which varies from ca. 40:1 to 150:1 (by weight) and depends on the phytoplankton group present and their nutritional status. Note that the Redfield Ratio, as in Eq 1, describes a reversible process, i.e. the respiration or decomposition of biomass. The break down of C will consume O2 and liberate N and P. A good example of such mass balance behavior with adverse effects is provided by blooms of the dinoflagellate genus Ceratium which sometimes result in hypoxic or anoxic events. Blooms of ceratian species can be stimulated by nutrient enrichment [57][187]. Hypoxic and anoxic ceratian blooms have been reported in the Kattegat [187], German Bight [130] and New York Bight [152], among other areas. During such events, the ceratian species respond in yield-dose behavior to available nutrients, with the population increasing until essential nutrients become exhausted. Nutrient exhaustion then prevents further growth and is even unable to support the existing biomass. The population becomes physiologically stressed; the cells loose their motility and sink to the bottom sediments where they rot. During their decomposition, O2 is utilized (Eq 1) which decreases in situ concentrations and pushes bottom water towards hypoxia or anoxia. Such occurrences and the degree of deoxygenation depend upon the amount of ceratian biomass being decomposed and the degree to which the bottom waters are ventilated by physical oceanographic processes (see [1]). Regions of bottom water stagnation, particularly during summer-autumn when ceratian blooms predominate, are predisposed to hypoxic blooms. Anoxic ceratian blooms are also facilitated by reduced grazing pressure on Ceratium species because their large cell size leads to predator-prey mismatches [340]. Anoxic events can accompany blooms of other dinoflagellate species [254][286], silicoflagellates (Fanuko, 1989), cyanobacteria [168], Phaeocystis [396], and diatoms [128]. However, it is often difficult to find a definitive link between the hypoxia (anoxia) developing during phytoplankton blooms and nutrient enrichment.
| | | 7.3 Eutrophication and Phytoplankton: the Mass Balance Approach |