| | 7.1 Introduction |
The role of nutrients in regulating the seasonal and regional abundance of phytoplankton, their blooms, species successions, and carbon production (primary production) is a classical problem in biological oceanography. The natural fertility of marine waters varies greatly because the amount of nutrients supplied through natural processes to support photosynthesis and growth of the phytoplankton varies regionally and seasonally. The Sargasso Sea, for example, is well known for its poor nutrient supply and oligotrophic nature, in contrast to the high productivity and fisheries yield of nutrient-rich upwelling regions. Nutrient enrichment of oligotrophic waters can benefit food chains, but coastal ecosystems, which are intrinsically productive because of natural nutrient recycling processes, can become degraded through excessive enrichment with anthropogenic nutrients [1][95][257]. Analyses and mitigation of the impact of eutrophication on coastal phytoplankton behaviour require an understanding of the photosynthesis - nutrient relationship. This coupled physiology regulates the basic, cellular responses of the phytoplankton to nutrient enrichment, while the population that develops is modified by food web processes. This physiology, and eutrophication as a process will be reviewed briefly before describing some specific responses of the phytoplankton to nutrient enrichment.
The phytoplankton (primary producers) require four primary inorganic macro-nutrients (NH4, NO3, PO4, SiO2) and five micro-nutrients (Fe, Cu, Zn, Mn, Mo) to fix carbon during photosynthesis. The photosynthesis equation can be written as an oxidation-reduction reaction having the general form:
2H2O + CO2 + Light -> (CH20) + H2O + O2
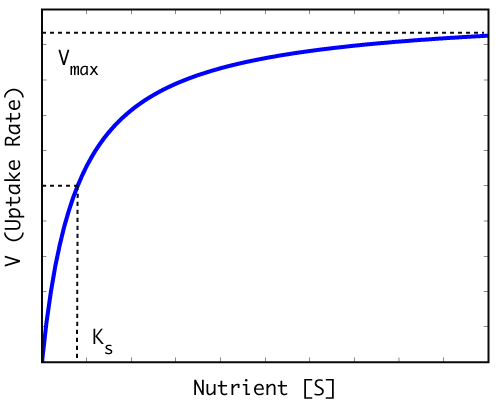
with the reaction mediated by the pigment chlorophyll a. Light energy is used to oxidize water yielding gaseous molecular oxygen [153]. The CO2 reduced during photosynthesis is temporarily fixed to an organic molecule (CH20) which serves as the substrate used in subsequent biosynthetic reactions to manufacture amino and nucleic acids, lipids, proteins, enzymes, etc. required for cellular growth and reproduction. This biosynthesis requires the concurrent assimilation of essential nutrients (nitrogen, phosphorus, micro-nutrients, etc.) during photosynthesis. The rate of nutrient assimilation is a function of its concentration, and increases hyperbolically with this (Figure 1). There are two kinetic characteristics of the nutrient uptake curve which vary among species and influence their responses to nutrient enrichment (or limitation) in competition with other species [432]: the maximum velocity of cellular uptake (Vmax) and the concentration of nutrient (Ks) at which uptake is one-half Vmax. The half-saturation constant, Ks, is particularly important since it determines the efficiency with which species take up nutrients at low concentrations. The higher the Ks constant, the less able the species in question to assimilate nutrients at low concentrations. On this basis, it might be expected that eutrophication would favor species less efficient in nutrient uptake.
Numerous experiments have shown that at increased rates of nutrient uptake cellular growth is stimulated, and an increase in population abundance results. Similar to the nutrient uptake curve (Figure 1), the population will increase up to an asymptotic level and remain relatively constant irrespective of further increases in nutrient concentration. The population at this upper (non-grazed) level corresponds to the carrying capacity for that particular species growing in response to the nutrient being supplied. The carrying capacity of a given nutrient level is not constant; it varies with nutrient type, concentration, accompanying growth factors, and among species. Excessively high nutrient levels, particularly NH4, can be inhibitory [464]. The amount of light (Eq. 1) available influences the effect of nutrients on photosynthesis at the cellular (individual) level, a relationship found also for natural populations [95]. Photosynthesis when deprived of light ceases, or decreases when nutrients are in short supply. During these conditions, the reverse reaction in Eq. 1 predominates or is favoured; i.e. oxygen is consumed (respiration) leading to the production of CO2. If the respiration pathway is prolonged and the ratio of photosynthetic oxygen production to respiration drops below 1.0, the decreasing oxygen level will push the water body towards hypoxia or, in extreme cases, to anoxia. Thus, nutrient loading can oxygenate and de-oxygenate a nutrient-enriched watermass [380].
In summary, the amount and types of nutrients available influence two coupled, cellular responses: the rate of nutrient uptake (Figure 1) and the rate of photosynthesis (Eq. 1). These responses, in turn, regulate the rates of cellular (species) growth and population growth (abundance). Eutrophication also has two distinct modes of ecosystem impact -- primary and secondary -- with the impacts of the nutrient disturbance further influenced by the other habitat conditions. The primary (direct) effect of nutrient enrichment is on the phytoplankton, both their abundance and species responses. This altered behavior impacts upper trophic level processes and components whose responses develop as secondary (indirect) effects of nutrification. Thus, both top-down (nutrient-phytoplankton responses) and bottom-up (grazer-phytoplankton responses) manipulations accompany nutrient enrichment of marine coastal systems. This complexity, the nutrient-photosynthesis relationship, the nine major macro- and micro-nutrients that regulate phytoplankton growth, and differences in species' nutritional physiology and kinetics make analysis of the influence of eutrophication on phytoplankton difficult. The following overview is therefore subject to modification as knowledge of eutrophication as a process and as a stress on marine ecosystems increases.
| | 7.1 Introduction |