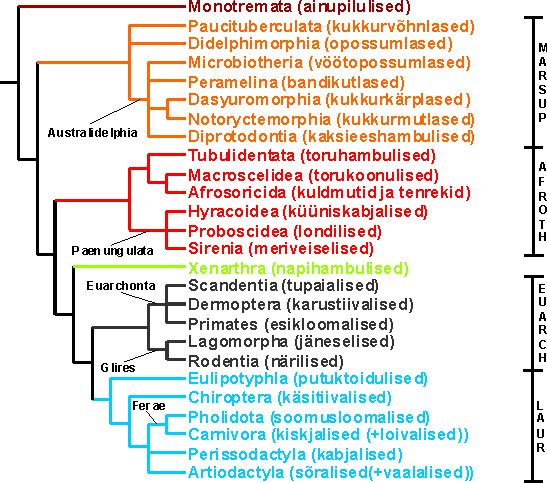
Joonis 3. Imetajate fülogenees Murphy et al. (2001b) (platsentaalid e. pärisimetajad), Amrine-Madsen et al. (2003) ja Baker et al. (2004) (kukrulised) järgi. Marsup - Marsupialia (kukrulised); Afroth - Afrotheria; Euarch - Euarchontoglires; Laur - Laurasiatheria. Euarchontoglires (=Supraprimates) ja Laurasiatheria ühendatakse klaadi Boreoeutheria (=Boreotheria).
Võib olla, et imetajate evolutsioon on peaaegu täielikult rekonstrueeritud (mis puutub tänapäeval elavatasse rühmadesse). Ei tohiks kahtlust olla, et kukrulised on sõsarrühmaks pärisimetajatele (Phillips & Penny, 2003) ja need mõlemad omakorda (i. e. Theria) sõsarrühmaks ainupilulistele. Loodetavasti on platsentaalide jaotamine nelja suurde rühma (Afrotheria, Xenarthra, Euarchontoglires, Laurasiatheria) samuti korrektne. Eksisteerivad siiski mõned vastuolud. Nimelt paljud uuringud on näidanud, et Euarchontoglires võib olla parafüleetiline teiste platsentaalide suhtes. Sellistele järeldustele on jõudnud enamik mitokondri genoomi järjestusel põhinevatest analüüsidest (e. g. Arnason et al., 2002). Kuid on avaldatud arvamust, et mitokondriaalne DNA evolutsioneerub liiga kiiresti lahendamaks imetajate varajast fülogeneesi (Corneli, 2002). Siiski on ka mõned rakutuuma geenidel põhinevad analüüsid jõudnud sarnastele järeldustele mitokondri genoomi omadega (Asher et al., 2003; Misawa & Janke, 2003). Samas jõudsid Reyes et al. (2004), kes lisasid analüüsidesse veel kaks närilise järjestust, mitokondri genoomi kasutades sarnastele järeldustele enamike tuumageenidega. Nende kõrvalekallete (e. g. Arnason et al., 2002; Asher et al., 2003; Misawa & Janke, 2003) põhjuseks on tõenäoliselt kas ebasobivate molekulaarsete fülogneesi meetodite ja mudelite kasutamine (õige molekuli järjestuse evolutsiooni mudeli kasutamine on oluline siis, kui mõned analüüsides kasutatavad taksonid näitavad ebatavalist nukleotiidset koostist või nukleotiidide asendumiskiirus on erinev võrreldes teistega) või väike taksoni valim (Springer et al., 2004). Hiljuti ilmunud ülevaade (Springer et al., 2004) on kenasti kokku võtnud platsentaalide molekulaarse fülogeneesi uuringud ning autorid on veendunud, et pärisimetajate evolutsioonipuu on praktiliselt rekonstrueeritud. Väitele, et imetajate fülogenees on seltsi tasemel peaaegu kindlaks tehtud, lisavad olulist toetust mobiilsete geneetiliste elementide (retrotransposoonide) analüüsid (Nishihara et al., 2005; 2006; Kriegs et al., 2006). Kõik peamised geenijärjestuste analüüside põhjal kindlaks tehtud uued klaadid (Afrotheria, Euarchontoglires, Laurasiatheria, Boreoeutheria) leidsid kinnitust, kui uuriti erinevate retrotransposoonide paiknemist pärisimetajate genoomides (Nishihara et al., 2005; 2006; Kriegs et al., 2006). Igat nimetatud nelja klaadi defineerib vähemalt 6-11 spetsiifilist retrotransposooni kindlas kromosoomi lookuses. Vastavalt nendele uutele tulemustele tuleks ülaltoodud joonist mõningates kohtades modifitseerida: Afrotheria sees tuleks kuldmutid ja tenrekid tõenäoliselt ühendada toruhambulistega (Nishihara et al., 2005), Xenarthra ei pruugi olla sõsarrühmaks Boreoeutheria klaadile (Kriegs et al., 2006) ning Laurasiatheria sees võivad käsitiivalised (Chiroptera) üllataval kombel olla sõsarrühmaks Pholidota+Carnivora+Perissodactyla klaadile (Nishihara et al., 2006). Käsitiivaliste (Chiroptera), kabjaliste (Perissodactyla) ja Ferae (soomusloomalised+kiskjalised) klaadile on pandud huvitav nimi: Pegasoferae (Pegasus on kreeka mütoloogias lendav hobune) (Nishihara et al., 2006).
Igatahes on paljude geenide (kümnete, sadade) järjestustel põhinevad fülogeneetilised analüüsid võimsad meetodid organismide evolutsiooni rekonstrueerimisel. Peale imetajate (Murphy et al., 2001b aheldasid analüüsimiseks näiteks 22 erineva geeni fragmenti; aluseks olid Madsen et al., 2001 ja Murphy et al., 2001a tööd) on sellised analüüsid häid tulemusi andnud ka mitmete teiste rühmade fülogeneesi väljaselgitamisel. Nagu näiteks Baldauf et al., 2000 (eukarüoodid); Bapteste et al., 2002 (amööbid); Rokas et al., 2003 (pärmseened). Rokas et al. (2003) (kes kasutasid rohkem kui saja geeni järjestusi) väitsid isegi, et neil on õnnestunud välja selgitada 8 pärmiliigi tõelised fülogeneetilised suhted. Kui analüüsiti üksikuid geene eraldi, siis saadi mitmeid omavahel vastuolus olevaid fülogeneesi puid. Aga kui analüüsi kaasati üheaegselt 106 geeni, jõuti olenemata analüüsimeetodist ainult ühele tulemusele, kusjuures saadud kladogrammi erinevate klaadide statistilised toetused olid maksimaalsed. Enam-vähem samadele tulemustele jõuti ka minimaalselt 20 geeni analüüsimisel. Sellest hoolimata ei saa lõpuni kindel olla, et 100 või kui tahes suure hulga geenide analüüsimisel jõutakse korrektse fülogeneesini. See niiöelda korrektne fülogenees võib mingil põhjusel siiski vale olla ja uute geenide lisamine analüüsi lihtsalt võimendaks viga. Sellisel juhul võib abiks olla hoopis taksonite, mitte molekulide, lisamine analüüsi (Hillis et al., 2003).
Helgen (2003) juhtis tähelepanu mõnedele omapärastele, väikestele ja vähetuntud imetajarühmadele, kelle fülogeneetiline positsioon vajaks veel selgitamist [nende hulgas olid ka piluhambulised (Solenodontidae)]. Roca et al. (2004) näitasid, et piluhambulised1 on Laurasiatheria klaadi kuuluvad kõige primitiivsemad putuktoidulised (Eulipotyphla).
Traditsiooniliselt on üheks kõige primitiivsemaks pärisimetajate rühmaks peetud putuktoidulisi. Shoshani & McKenna (1998) järgi on napihambulised (Xenarthra) sõsarrühmaks ülejäänud platsentaalidele (i. e. Epitheria). Epitheria jaguneb omakorda kaheks: putuktoidulised (Lipotyphla) ja ülejäänud. Mõlemad molekulaarsete andmete järgi defineeritud klaadid - Eulipotyphla ja Afrosoricida - kuulusid Shoshani & McKenna (1998) järgi Lipotyphla hulka. Siiski möönsid nad, et see putuktoiduliste rühm võib olla polüfüleetiline. Kui ülal toodud fülogeneesi puu on korrektne, siis tähendab see seda, et putuktoidulised laias mõttes on difüleetiline rühmitus. Varem on putuktoiduliste hulka arvatud ka torukoonulisi (Macroscelidea), nii ka teatmeteoses Loomade elu (Poots, 1987). Putuktoidulisi on peetud kõige esimesteks ja vanimateks pärisimetajateks sellepärast, et fossiilsete leidude järgi olidki praktiliselt kõik pärisimetajad Kriidi ajastul sellised (väiksed, varjatud ja öise eluviisiga; nagu tänapäeva putuktoidulised). Samas näitasid Springer et al. (2003), et enamike pärisimetajate seltside teke jääb Kriidi ajastusse (u. 80-60 miljonit aastat tagasi; krooni rühma e. tänapäeval elavate platsentaalide ühine eellane jääb ~110 milj. aasta tagusesse aega), mis tähendab et putuktoiduliste kõrval pidid dinosauruste ajal ringi liikuma ka kiskjalised, esikloomalised, närilised ja teised. Ilmselt olidki enamus imetajaid Kriidi ajastul putuktoiduliste laadsed. Tänapäeval elavate seltside esindajad pärinevad lihtsalt Kriidi ajastul elanud erinevatest "putuktoiduliste" rühmadest. Pärast dinosauruste (ja enamike teiste elusorganismide) väljasuremist 65 miljonit aastat tagasi toimus imetajate adaptiivne radiatsioon (mitmekesistumine). Erinevad "putuktoiduliste" rühmad hõivasid dinosauruste poolt vabaks jäetud erinevad ökonišid ja nii kujunesid üksteisest hästi eristatavad tänapäeva imetajate seltsid. Ainult üksikud putuktoidulised ei muutunud välimuselt eriti pärast dinosauruste väljasuremist, jäädes truuks oma vanale eluviisile. Nendeks on siis kahes erinevas pärisimetajate klaadis paiknevad Eulipotyphla (Laurasia mandri putuktoidulised) ning Afrosoricida ja Macroscelidea (Aafrika mandri putuktoidulised).
1 - Loomade elu (Poots, 1987) järgi on selle putuktoiduliste sugukonna nimi pilukoonlased, kuid eesti terioloogi A. Miljutini soovituse kohaselt võiks kasutada eesti keeles nimetust piluhambulised, kuna see on korrektsem tõlge teaduslikust nimest.
Viited
Amrine-Madsen, H. Scally, M. Westerman, M., Stanhope, M. J., Krajewski, C., Springer, M. S. 2003. Nuclear gene sequences provide evidence for the monophyly of australidelphian marsupials. Molecular Phylogenetics and Evolution 28: 186-196.
Arnason, U., Adegoke, J. A., Bodin, K., Born, E. W., Esa, Y. B., Gullberg, A., Nilsson, M., Short, R. V., Xu, X & Janke, A. 2002. Mammalian mitogenomic relationships and the root of the eutherian tree. Proceedings of National Academy of Science. USA 99: 8151-8156. [pdf]
Asher, R. J., Novacek, M. J., & Geisler, J. H. 2003. Relationships of Endemic African Mammals and Their Fossil Relatives Based on Morphological and Molecular Evidence. Journal of Mammalian Evolution 10: 131-194.
Baker, M. L., Wares, J. P., Harrison, G. A. & Miller, R. D. 2004. Relationships among the Families and Orders of Marsupials and the Major Mammalian Lineages Based on Recombination Activating Gene-1.
Journal of Mammalian Evolution 11: 1-16.Baldauf, S. L., Roger, A. J., Wenk-Siefert, I., Doolittle, W. F. 2000. A Kingdom-Level Phylogeny of Eukaryotes Based on Combined Protein Data. Science 290: 972-977.
Bapteste, E., Brinkmann, H., Lee, J. A., Moore, D. V., Sensen, C. W., Gordon, P., Duruflé, L., Gaasterland, T., Lopez, P., Müller, M. & Philippe, H. 2002. The analysis of 100 genes supports the grouping of three highly divergent amoebae: Dictyostelium, Entamoeba, and Mastigamoeba. Proceedings of the National Academy of Sciences. USA 99: 1414–1419. [pdf]
Corneli, P. S. 2002. Complete Mitochondrial Genomes and Eutherian Evolution.
Journal of Mammalian Evolution 9: 281-305.Helgen, K. M. 2003. Major mammalian clades: a review under consideration of molecular and palaeontological evidence. Mammalian Biology 68: 1-15.
Hillis, D. M., Pollock, D. D., McGuire, J.A. & Zwickl, D. J. 2003. Is sparse taxon sampling a problem for phylogenetic inference? Systematic Biology 52: 124-126.
Kriegs, J. O., Churakov, G., Kiefmann, M., Jordan, U., Brosius, J. & Schmitz, J. 2006. Retroposed Elements as Archives for the Evolutionary History of Placental Mammals. PLoS Biology 4: e91. [pdf]
Madsen O., Scally, M., Douady, C.J., Kao, D.J., DeBry, R.W., Adkins, R., Amrine, H.M., Stanhope, M.J., de Jong, W.W., Springer, M.S. 2001. Parallel adaptive radiations in two major clades of placental mammals. Nature 409: 610-614.
Misawa, K. & Janke, A. 2003. Revisiting the Glires concept—phylogenetic analysis of nuclear sequences. Molecular Phylogenetics and Evolution 28: 320–327.
Murphy, W.J., Eizirik, E., Johnson, W.E., Zhang, Y.P., Ryder, O.A. & O'Brien, S.J. 2001a. Molecular phylogenetics and the origins of placental mammals. Nature 409: 614-618.
Murphy, W. J., Eizirik, E., O'Brien, S. J., Madsen, O., Scally, M., Douady, C. J., Teeling, E., Ryder, O. A., Stanhope, M. J., de Jong, W. W. & Springer, M. S. 2001b. Resolution of the Early Placental Mammal Radiation Using Bayesian Phylogenetics. Science 294: 2348-2351.
Nishihara, H., Satta, Y., Nikaido, M., Thewissen, J. G. M., Stanhope, M. J. & Okada, N. 2005. A Retroposon Analysis of Afrotherian Phylogeny. Molecular Biology and Evolution 22: 1823–1833. [pdf]
Nishihara, H., Hasegawa, M. & Okada, N. 2006. Pegasoferae, an unexpected mammalian clade revealed by tracking ancient retroposon insertions. Proceedings of the National Academy of Sciences. USA 103: 9929–9934. [pdf]
Phillips, M. J. & Penny, D. 2003. The root of the mammalian tree inferred from whole mitochondrial genomes. Molecular Phylogenetics and Evolution 28: 171-185.
Poots, L. 1987. Loomade elu. 7. köide. Imetajad. Valgus, Tallinn.
Reyes, A., Gissi, C., Catzeflis, F., Nevo, E., Pesole, G. & Saccone, C. 2004. Congruent Mammalian Trees from Mitochondrial and Nuclear Genes Using Bayesian Methods. Molecular Biology and Evolution 21: 397-403. [pdf]
Roca, A. L., Bar-Gal, G. K., Eizirik, E., Helgen, K. M., Maria, R., Springer, M. S., O'Brien, S. J. & Murphy, W. J. 2004. Mesozoic origin for West Indian insectivores. Nature 429: 649-651.
Rokas, A., Williams, B. L., King, N. & Carroll., S. B. 2003. Genome-scale approaches to resolving incongruence in molecular phylogenies. Nature 425: 798-804.
Shoshani, J. & McKenna, M. C. 1998. Higher Taxonomic Relationships among Extant Mammals Based on Morphology, with Selected Comparisons of Results from Molecular Data. Molecular Phylogenetics and Evolution 9: 572-584.
Springer, M. S., Murphy, W. J., Eizirik, E. & O'Brien, S. J. 2003. Placental mammal diversification and the Cretaceous-Tertiary boundary. Proceedings of National Academy of Science. USA 100: 1056-1061. [pdf]
Springer, M. S., Stanhope, M. J., Madsen, O. & de Jong, W. W. 2004. Molecules consolidate the placental mammal tree. Trends in Ecology & Evolution 19: 430-438.