SUGUDE SUHE
© Tiit Teder 2000
Sissejuhatus
Fisher'i teooria
Hamilton'i teooria
Trivers-Willard'i teooria
Geneetiline kontroll
Sugude suhe erinevates loomarühmades
Sugude suhe parasiitsetel kiletiivalistel
Sugude suhe inimesel
Sugude suhet määravad füsioloogilised mehhanismid imetajatel
Kasutatud kirjandus
Teemakohast internetist
Kordamisküsimused
SISSEJUHATUS
Sugude suhte uurimine on evolutsioonilise bioloogia üks kőige edukamaid harusid. Aines omab detailset teoreetilist pőhjendatust ning paljud teooria ennustused on leidnud ka kinnitust vaatluste ja eksperimentide näol. Sugude suhte probleemist räägib oma käitumisökoloogia loengutes ka prof. Raivo Mänd (vt. http://www.loodus.ee/kaitumisokoloogia/ko8.html).
Sugude suhte probleemistik algab sellest, et vanemad peavad jagama sigimispingutust poegade ja tütarde vahel (nüüd ja edaspidi: kui sugude suhet on kuskil väljendatud kvantitatiivselt, siis peetakse silmas isaste/emaste suhet, mitte vastupidi). Vőtame näiteks lokaalpopulatsiooni, kus on 100 isast ja 10 emast. Oletame veel lihtsuse mőttes, et nii isased kui emased paarituvad vaid ühe korra. See tähendab, et 90% isastest jääb paaritumata. Sellisel puhul saavad eelise emane, kes produtseerivad tütreid, kuna tütreid produtseerides kasvab tema kohasus kiiremini. Tasakaal taastub siis, kui tütreid ja poegi produtseeritakse vőrdsel arvul. Seega toimub sagedusest sőltuv valik (frequency-dependent selection), kuna arvukam vorm - antud juhul isased - on vähemedukas kui haruldane vorm.
Sugude suhte teooria ajalugu ulatub Darwinini, kes leidis, et sugude suhe vőiks olla üks omadus, millele looduslik valik toimib, küll aga ei osanud ta välja pakkuda teooriat, miks sugude suhe on tavaliselt lähedane 1:1-le. Tsiteerides Darwinit: "... but I now see the whole problem [of sex ratio] as so intricate that it is safer to leave its solution for the future". Esimese korralikult argumenteeritud selgituse sellele pakkus välja Fisher (1930, rf. Charnov 1982, Godfray 1994), kelle teooria on aluseks olnud ka kőigile peamistele sugude suhte teooria edasiarendustele.
FISHER'I TEOORIA (1930)
1) Oletame, et poegade ja tütarde produtseerimine on vőrdselt kulukas. Emase kohasus (fitness) vőrdub sel juhul tütarde arvuga, millele on liidetud poegade poolt viljastatud emaste arv.
a) Kui populatsioonis on emaste-isaste suhe 1:1, siis üks poeg viljastab keskmiselt ühe emase, seega on poja ja tütre osa emase kohasusse vőrdne.
b) Kui populatsioonis on isaseid rohkem kui emaseid, siis üks poeg viljastab keskmiselt vähem kui ühe emase (mőni viljastab rohkem, mőni mitte ühtegi) ja on emale seega suhteliselt vähem väärtuslik kui tütar. Iga mutantne geen, mille kandja produtseerib sellistes tingimustes rohkem tütreid kui poegi, on loodusliku valiku poolt soositud ning selliste sagedus populatsioonis kasvab, pőhjustades 1:1 sugude suhte taastumise.
c) Kui populatsioonis on emaseid rohkem kui isaseid, käivitub teooria kohaselt vastupidine protsess, mille tulemusena jällegi taastub 1:1 sugude suhe.
d) Ainuke sugude suhe, mille puhul valik toimib mutantsete geenide vastu, ongi 1:1 sugude suhe.
Fisher'i teoorias on seega liikumapanevaks jőuks sagedusest sőltuv valik, mis tőukab populatsiooni olekusse, kus sugude suhe on 1:1.
2) Kui poegade ja tütarde produtseerimine ei ole vőrdselt kulukas, siis tuleb emase kohasust mőőta järglaskonda paigutatud ressursi ühiku kohta. Valik soosib geene, mis suurendavad selle sugupoole produktsiooni, mille mőju kohasusele on ressursi ühiku kohta suurem.
Et illustreerida seda oletame, et ühe tütre produtseerimiseks läheb 2 korda rohkem energiat kui ühe poja produtseerimiseks. Edasi oletame, et üks tütar suurendab emase kohasust 1 ühiku vőrra, üks poeg aga 0,6 ühiku vőrra. Seega ühe energiaühiku kulutades poegade peale suureneb emase kohasus 1,2 ühiku vőrra, ühe energiaühiku kulutamisel tütarde peale aga suureneb emase kohasus vaid 1 ühiku vőrra. Niisiis, kuigi üks tütar suurendab emase kohasust rohkem kui üks poeg, soosib valik antud juhul poegade enamusega sugude suhet, kuna tütre produtseerimise hind on poja produtseerimisest märksa kőrgem. Ainuke stabiilne sugude suhe tekib poegadesse ja tütardesse paigutatud ressursside vőrdsuse korral.
Sugude erinev suremus ei avalda mőju ei vanemlooma kohasusele paigutatud ressursi ühiku kohta ega sündinute sugude suhtele. Näiteks, kui 50% poegadest surevad enne reproduktiivsesse ikka jőudmist, tütred aga jäävad ellu, siis iga poeg paaritub keskmiselt kahe emasega. Vanemlooma kohasus seejuures ei muutu: ühest küljest vanemlooma kohasus poegade kőrge suremuse tőttu küll langeb, teisest küljest on igal ellujäänul aga vőimalik paarituda keskmiselt kahe emasega, mis sisuliselt taastab vanemlooma kohasuse. (Vanemlooma poolt produtseeritud järglaste sugude suhet nimetatakse primaarseks (primary sex ratio), järglaskonna reproduktiivsesse ikka jőudnud isendite sugude suhet sekundaarseks (secondary sex ratio) ehk operatiivseks (operational sex ratio).)
Fisher'i teooria nőuab mitmete eelduste täidetust.
(1) Paaritumine peab olema panmiktiline, st. ühe vanemlooma järglaskond ei tohiks omavahel paarituda. Mittekattuvate pőlvkondade puhul peaks toimuma enne uut paarumishooaega segunemine.
(2) Kohasus peaks kasvama lineaarselt ressursside paigutusega kummassegi sugupoolde. See tähendab, et kui emane paigutab ühte sugupoolde kaks korda rohkem ressursse, siis tema kohasus peaks tőusma ka kaks korda.
(3) Emaslooma geneetiline sugulus oma poegade ja tütardega peab olema ühesugune. Tavaliselt diploidsetel loomadel see nii ka on (eranditest vt. allpool). Oletame, et emalooma ning tema tütarde sugulus on suurem kui emalooma ja poegade sugulus. Sel juhul on mingi emalooma geeni sattumine järgmisesse pőlvkonda läbi emasliini tőenäolisem, mis kokkuvőttes viib emaste enamusega sugude suhte kujunemiseni.
Shaw & Mohler (1953, rf. Charnov 1982) arendasid Fisheri teooriat edasi rakendades ESS (evolutsiooniliselt stabiilne strateegia: mingi tunnuse väärtus tasakaalulises olekus) lähenemist (ESS mőiste tuli kasutusele Maynard-Smith'i (1976) tööst).
Oletame, et meil on mittekattuvate pőlvkondadega populatsioon, mis koosneb isenditest, kellel on mingi tunnuse väärtuseks Z (näiteks sugude suhe). Populatsiooni tuuakse haruldane genotüüp, millel on tunnuse alternatiivne väärtus Z'. Kui eksisteerib mingi Z väärtus, mille korral valik toimib tunnuse kőigi alternatiivsete väärtuste vastu, siis nimetatakse seda tunnuse väärtust ESS'ks. Shaw & Mohler näitasid, et mutantse geeni levimine populatsioonis sőltub tema kohasusest tavalise tüübi suhtes. Mutandi kohasuse vőrrand on järgmine:

Seos on ehk paremini arusaadav, kui kirjutada see ümber järgmisele kujule:
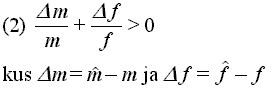
Sőnastatult vőiks see vőrrand kőlada järgmiselt: valik soosib sugude suhet muutvat mutantset geeni siis, kui vőit kohasuses läbi ühe sugupoole ületab kaotuse läbi teise sugupoole. Selle matemaatilist kuju nimetatakse Shaw-Mohler'i vőrrandiks.
Seega vőib lühidalt öelda, et valik maksimiseerib suurust
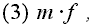
kus m on kohasus läbi isasfunktsiooni ja f on kohasus läbi emasfunktsiooni. See maksimiseeritav suurus on ESS sugude suhe.
HAMILTON'I TEOORIA (1967)
Fisher'i teooria eeldas juhuslikku paaritumist. Hamilton (1967) oli esimene, kes näitas, et 1:1 ei pruugi olla ainuke vőimalik ESS juhul, kui Fisher'i panmiksise (populatsiooni liikmed paarituvad omavahel juhuslikult) eeldust lődvendada.
Oletame, et meil on laigulises keskkonnas elav liik. Iga laigu hőivavad n paarunud emast, kes produtseerivad seal vőrdse arvu järglasi. Need arenevad üheskoos ning lőpuks paarituvad omavahel. Uue pőlvkonna emased hajuvad seejärel laiali ja tekivad uued juhuslikud n emasest koosnevad grupid.
Hamilton näitas, et sellise populatsiooni struktuuri korral tekib emaste enamusega sugude suhe. ESS sugude suhe on sel juhul (n-1)/2n, kus n on emaste arv laigul. Kui ühe laigu hőivab vaid üks emane, peaks sugude suhe olema 0. Hamiltoni seletuse järgi sellisel juhul emane muneb ainult niipalju isaseid, et need viljastaksid kőik tema tütred. Kui emaste arv laigul kasvab, siis paaritumine toimub suure osa populatsiooni liikmete vahel (panmiksis) ning sugude suhe läheneb Fisher'i 1:1-le, nagu vőib ennustada juhu jaoks, kui paaritumine toimub juhuslikult. Hamilton nimetas käsitletud emaste enamusega sugude suhteni viivat mehhanismi lokaalseks paarumiskonkurentsiks.
Levinuim seletus Hamiltoni mudeli emaste enamusega sugude suhetele on järgmine. Kui ühest soost järglased konkureerivad omavahel rohkem kui teisest soost järglased, siis ESS sugude suhe on selline, kus enamuses on see sugupool, kes konkureerib omavahel vähem paarumisvőimaluste pärast. Lokaalse paarumiskonkurentsi puhul on vähemalt kaks pőhjust, miks emaste enamusega sugude suhe on vanemloomale soodus.
1) Vähem poegi tähendab poegade vahel madalamat konkurentsitaset partnerite pärast.
2) Rohkem tütreid tähendab rohkem potentsiaalseid partnereid poegadele.
Sugude suhte edasisele kahanemisele seab piirid sagedusest sőltuv valik.
TRIVERS-WILLARD'I TEOORIA (1973)
Trivers & Willard (1973) täiendasid sugude suhte teooriat edasi, uurides, kuidas sugude suhted on seotud vanemate konditsiooniga. Teisisőnu täiendasid nad teooriat mitte sellega, milline on populatsiooni üldine sugude suhe, vaid kuidas see varieerub populatsiooni liikmetel sőltuvalt nende konditsioonist.
Oletame, et meil on imetajate populatsioon, mille emased varieeruvad oma füsioloogiliselt konditsioonilt. Emase konditsioon omakorda mőjutab järglaste sigimisvőimekust, s.t. kehvas konditsioonis emaste pojad ja tütred on ka ise kehvemad sigijad, samal ajal kui heas konditsioonis emaste järglased on üle keskmise vőimekad. Seega vőidavad nii pojad kui tütred, kui neil on heas konditsioonis ema.
Trivers'i & Willard'i argumentatsiooni järgi imetajatel pojad vőidavad aga heast konditsioonist rohkem kui tütred, kuna isaste sigimisvőimekus on tugevamini korreleeritud konditsiooniga teiste isaste suhtes. Kui isaste puhul vőib kehv konditsioon sigimisvőimalused kahandada nullini, siis emaste puhul on selline vőimalus harv. Seega isaste puhul konditsiooni kasvades sigimisedukus suureneb kiiremini kui emastel. Sellest nad järeldasid, et heas konditsioonis emased peaks produtseerima rohkem poegi, kehvas konditsioonis emased rohkem tütreid.
GENEETILINE KONTROLL
Oluline täiendus sugude suhte teooriale oli geneetilise kontrolli osa tähtsustamine sugude suhte mőjutamisel (Lewis 1941, rf. Frank 1990; Hamilton 1967; Godfray & Werren 1996). Varasemates sugude suhteid käsitlevates töödes eeldati, et soo määravad rakutuuma geneetilised elemendid, eelkőige kromosoomid, mis päranduvad vőrdselt nii poegadele kui tütardele. Lisaks sellele on organismis aga veel teisi geneetilisi elemente, mille pärandumine ei ole sümmeetriline, s.t. elemendid, mis päranduvad ühele sugupoolele, aga teisele mitte. Asümmeetria geneetiliste elementide pärandumisel toob kaasa vähemasti kaks olulist tagajärge:
1) valik viib sugude suhte kaldumisele selle sugupoole suunas, kes selliseid elemente pärandab, (reeglina on selleks emane);
2) genoomi erinevate elementide vahel on geneetiline konflikt soo paigutamisel.
Oletame, et kontroll sugude suhte üle on emasliini pidi edasikantavate tsütoplasmaatiliselt päranduvate geneetiliste elementide käes. Kuna nende geenide edukus sőltub ainult tütardest, siis on edukam see tsütoplasmaatiline geneetiline element, mille kandja produtseerib vaid tütreid. Kuna rakutuuma geenidele toimib valik, mis soosib enam-vähem vőrdset ressursside paigutamist erinevatesse sugupooltesse, siis tekib erinevate geneetiliste elementide vahel konflikt. Evolutsiooniline vőidujooks nende elementide vahel on oluliseks teguriks sugude suhet evolutsioonis.
Sarnane on olukord aga ka imetajate ja lindude puhul, kelle sugude suhet kontrollivad elemendid on sugukromosoomides. Imetajatel on teatavasti XX emased ning XY isased. Kui me oletame, et sugude suhet kontrollivad geenid on Y-kromosoomis, siis peaks tekkima tugevalt isaste enamusega sugude suhted, kuna selliste geenide levimine populatsioonis sőltub ainult isastest, s.t. Y-kromosoomi suhtes isad ja tütred ei ole sugulased.
Lindudel on vastupidi, neil on ZZ isased ja WZ emased. Järelikult, kui sugukromosoomidel on geneetiline kontroll sugude suhte üle, peaks lindudel kujunema emaste enamusega sugude suhted.
SUGUDE SUHE ERINEVATES LOOMARÜHMADES
Sugude suhe parasiitsetel kiletiivalistel
Üks kőige pőhjalikumalt sugude suhte osas uuritud loomarühmi on parasiitsed kiletiivalised, (käguvaablased, juuluklased jne. - vt. pilt käguvaablasest). Hamilton (1967) esitades oma lokaalse paarumiskonkurentsi hüpoteesi pidas silmas just parasitoide, kui eriti sobivat loomarühma oma hüpoteesi testimiseks. Nimelt on parasiitsetel kiletiivalistel nii geneetiline kui ökoloogiline eelsoodumus emaste enamusega sugude suhete esinemisele lokaalse paarumiskonkurentsi tőttu. Ühest küljest, nagu enamus kiletiivalisi, on nad geneetiliselt haplodiploidsed (haplodiploidsetel liikidel on isased haploidsed, emased diploidsed) s.t. isased arenevad viljastamata munarakkudest, emased viljastatud munarakkudest. Seega on munetavate järglaste sugude suhe otseselt emaslooma kontrolli all. Teisest küljest, ökoloogiliselt esineb vendade vahel sageli paarumiskonkurents. Eriti sage on see gregaar- ehk karjaparasitoidide (ühest peremeesorganismist toitub palju arenevaid parasitoide) puhul ning karjaviisiliselt elavaid peremehi ründavate parasitoidide puhul. Karjaparasitoidide järglased olles lőpetanud peremehest toitumise, nukkuvad selle vahetus läheduses. Isased valmikud kooruvad enam-vähem sünkroonselt nii omavahel kui sünkroonis sama järglaskonna emastega. Seega on tőenäoline nii ődede-vendade omavaheline sigimine kui vendade vaheline paarumiskonkurents ődede pärast. Sama juhtub siis, kui üksik- ehk solitaarparasitoid ründab karjaviisiliselt toituvaid peremehi. Siia kuuluvad näiteks paljud munaparasitoidid (vt. munaparasitoidi pilti, vőib suhteliselt aeglaselt tulla, kuna link on Ameerikasse), samuti diskreetsetel laikudel (näiteks seen) toituvate liikide parasitoidid.
Lokaalse paarumiskonkurentsi hüpotees teeb rea testitavaid ennustusi.
a) Kui vőrrelda erinevaid lähedasi liike, siis peaks teooria järgi sugude suhe (isased/emased) olema seda väiksem, mida tugevam on lokaalne paarumiskonkurents. Waage (1982) testis seda ennustust ühe munaparasitoidide sugukonna (Scelionidae) näitel. Osa parasitoidiliike paigutavad oma järglased suurtesse peremeesliigi munakogumitesse, teised väiksematesse munakogumitesse, osa liike aga üksikult paiknevatesse peremeesliigi munadesse. Waage oletas, et väikeste munakogumite puhul parasiteeritakse enamus mune ühe parasitoidid poolt, seega on oodatav paarumiskonkurents teravam, kui suurte munakogumite puhul. Teisest küljest ei ole paarumiskonkurentsi oodata ka üksikutesse munadesse paigutatavate järglaste vahel, kuna need on üksteisest selleks liiga kaugel. Uurimuse tulemused kinnitasid seda loogikat: sugude suhe oli kőige suurem suurtesse munakogumitesse ning üksikmunadesse munenud parasitoidide järglaskonnas, suhteliselt väiksem aga väikestes munakogumites.
b) Kui emase järglaskonnas toimub poegadevaheline konkurents, siis hüpoteesi järgi peaks emased produtseerima vaid nii palju poegi, kui palju on neid vaja tütarde paaritamiseks. Seega karjaparasitoidide puhul peaks sugude suhe sőltuma emase poolt munetud pesakonna suurusest (clutch size): mida suurem pesakond, seda suhteliselt vähem peaks olema poegi. Griffiths & Godfray (1988) uurisid seda kääbusherilaste (Bethylidae) sugukonna näitel. Tulemused kinnitasid seda hüpoteesi: suuremates pesakondades oli isaste osakaal väiksem. Samas peab nende tulemuste puhul, kus vőrreldakse erinevaid liike uurimata nende fülogeneesi, jääma siiski veidi ettevaatlikuks. Liigid on üksteisega fülogeneetiliselt erineval määral seotud ning neid ei ole analüüsides korrektne vaadelda iseseisvate punktidena. Kummagi siinkäsitletud töö puhul ei ole seda tehtud.
c) Hamiltoni mudel ennustab, et antud parasitoidi sugude suhet mőjutab teiste emaste arv uuritaval laigul. Ennustust on vőimalik testida manipuleerides emaste arvuga mingil laigul. Seejuures eeldatakse, et emane on vőimeline hindama teiste emaste arvu laigul ning reguleerima vastavalt sellele järglaste sugude suhet. Esimesena testis seda ennustust Werren (1983) nukukireslastel (Pteromalidae). Werren näitas, et need emased, kes olid laigul üksi, produtseerisid äärmusliku emaste enamusega järglaskonna, emaste arvu kasvades laigul, toimus ka sugude suhte suurenemine, s.t. emased produtseerisid poegi suhteliselt rohkem.
Sugude suhe solitaar- ehk üksikparasitoididel
On korduvalt näidatud, et kui peremehe populatsioonis isendite kehasuurus varieerub, siis on tendents sellele, et parasitoidid paigutavad tütreid suurematesse ning poegi väiksematesse peremeesloomadesse. On teada, et reeglina on emased parasitoidid suuremad kui isased (suguline dimorfism!!!) ning suuremad emased on reeglina viljakamad kui väikesed. Isaste puhul ei mängi kehasuurus parasitoididel reeglina niisuurt rolli, kuigi ka suuremad isased on tavaliselt mőnevőrra edukamad emastega paarumisel.
Charnov (1979) modelleeris sellist süsteemi ning näitas, et niisugustel tingimustel toimib valik emastele tőepoolest nii, et need paigutaksid tütreid suurematesse ning poegi väiksematesse peremeesisenditesse. Seejuures ei ole peremehe kehasuuruse mőiste siin absoluutne, vaid sőltub antud kehasuuruse jaotusest peremehel. See, millises vahekorras tütreid ja poegi erineva suurusega peremeesisenditesse munetakse, sőltub ka väikeste ja suurte peremeesisendite suhtest populatsioonis. Kui väikseid on palju, munetakse neisse kőik pojad ja ka osa tütreid. Vastupidisel juhul - kui suuri peremeesisendeid on palju - munetakse neisse kőik tütred ja ka osa poegi.
Muud sugude suhet mőjutavad faktorid parasiitsetel kiletiivalistel (King 1987; Hardy 1992, 1994)
Kuna parasiitsetel kiletiivalistel on sugude suhet uuritud väga laialdaselt, siis teen ülevaate, mis peale peremehe suuruse vőib mőjutada sugude suhet.
A. Vanemlikud omadused
1) Kui emaste koorumise ja sellele järgneva paaritumise vaheline aeg on pikk, siis see vőib viidata isaste vähesusele. Valik peaks sel juhul soosima emaseid, kes produtseerivad rohkem poegi.
2) Mida elu lőpu poole, seda rohkem poegi emased produtseerivad. Arvatakse, et selle pőhjuseks vőivad olla seemnerakkude ammendumisest vői seemnerakkude eluea pikkusest tulenevad efektid.
3) Emane vőib ka peale paaritumist muneda rohkem isaseid, kui spermidel kulub teatud aeg viljastumispaika jőudmiseks.
B. Keskkonnatingimused
4) Kőrgetel ja madalatel temperatuuridel on sugude suhted reeglina suure poegade enamusega. Ekstreemsed temperatuurid vőivad sugude suhet mőjutada järgmiselt: a) steriliseerida isased; b) pärssida liikumist ja sellega takistada paaritumist.
C. Peremehe omadused
5) Läbi kehasuuruse vőivad sugude suhtele mőju avaldada ka peremehe vanus (positiivses korrelatsioonis kehasuurusega), sugu (emased on peremeesliikidel tavaliselt suuremad) ja liik (osa liike suuremad, osa väiksemad).
Sugude suhe inimesel
Et see sugude suhte asi kaugeks ei jääks, tuleb järgnevalt juttu Soomes tehtud tööst, mis käsitleb sugude suhet inimesel (Lummaa et al. 1998). Kőigepealt kordamiseks veel kaks eespool defineeritud mőistet, mida siin kasutan. Operatiivse sugude suhte all peetakse silmas reproduktiivses eas olevate isaste ja emaste suhet populatsioonis. Sündinute sugude suhe mőiste sisu on ilmselt niigi arusaadav.
Antud töös kasutati vanade kirikuraamatute andmeid, kust oli vőimalik teada saada reproduktiivses eas (15-50 aastat) meeste ja naiste arv kihelkonniti, vastsündinute sugude suhe ja sündimus kihelkonnas. Uuriti andmeid Soome 21 kihelkonna kohta, mis paiknesid Soome erinevates piirkondades erinevates keskkonnatingimustes. Uurimus kattis ajavahemikku 1775-1850, mil industrialiseerimine ning tervishoid olid veel nőrgal järjel ning avaldasid vähest mőju elustandarditele ning ellujäämusele. Migratsioon erinevate kihelkondade vahel oli madal ning enamus abieludest toimus ühe kihelkonna piires.
Töös uuriti, milline on operatiivse sugude suhte ning sündinute sugude suhte vahekord. Ennustati, et kui operatiivne sugude suhe on emaste enamusega (st. reproduktiivses eas emaseid on populatsioonis rohkem kui isaseid), siis poegi sünnib suhteliselt rohkem kui tütreid. Lisaks sellele ennustati, et 1:1 operatiivne sugude suhe peaks maksimiseerima sündimuse ja viima sellega populatsiooni arvukuse kasvuni. Seda sellepärast, et 1:1 operatiivse sugude suhte puhul on teoreetiliselt populatsiooni igal isendil vastassoost partner, nihkes sugude suhte puhul aga jääb osa indiviide sigimisest kőrvale partneri puudumise tőttu (eeldusel, et tegemist on monogaamse liigiga, mida inimese kohta vőib teatud mööndustega ka öelda).
Töö tulemused kinnitasid ennustusi.
1) Operatiivne sugude suhe mőjutas sündinute sugude suhet. 21-st kihelkonnast 14-s viis emaste enamusega operatiivne sugude suhe isaste enamusega sugude suhtele sündinute puhul.
2) Sündimus suurenes operatiivse sugude suhte suurenedes. Nii oli see 21-st 15-s kihelkonnas.
NB! See, et valik soosib haruldasema sugupoole produktsiooni, on teada ka maailmasődade järgsetest kogemustest, kui toimus lühiajaline, kuid selge meeste sündimuse kasv vőrreldes naistega.
Sugude suhet määravad füsioloogilised mehhanismid imetajatel
Eelpool käsitletud haplodiploidsete kiletiivaliste puhul on selge, et emane saab sugude suhet käitumuslikult reguleerida. Diploidsetel organismidel, nende hulgas imetajatel ja lindudel, on vastavad mehhanismid aga seniajani vähetuntud. Imetajate puhul on küllaldaselt informatsiooni füsioloogiliste mehhanismide kohta, mis teoreetiliselt vőiksid mängida olulist rolli sündinute sugude suhte määramisel. Ülevaate vőimalikest sugude suhet reguleerivatest mehhanismidest on teinud Krackow (1995), millest siinkohal esitan lühikese kokkuvőtte.
a) Spermi valik
Spermi valik eeldab, et
• X- ja Y-kromosoomi kandvate seemnerakkude vőime jőuda munarakuni erineb; vői
• tőenäosus munarakku viljastada on sőltuvalt spermi tüübist (X vői Y) erinev.
Esimese mehhanismi järgi vőiks toimuda sugude suhte regulatsioon hamstritel, kelle puhul on näidatud sugude suhte sőltuvust suguteede pH-st. Kuna spermide liikuvus sőltub pH-st ning see erineb X- ja Y-kromosoomi kandvatel spermidel, siis vőib see viia eri tüüpi seemnerakkude erineva tiheduseni viljastamispaigas. Ka inimese puhul on teada, et X- ja Y-kromosoomi kandvad spermid erinevad oma liikuvuselt ning sőltub pH-st. Teist mehhanismi on seostatud eelkőige elektriliste nähtustega, näiteks erinevat tüüpi spermide erinevate pinnalaengutega.
b) Embrüo ja emaka arengu sünkroonsus
Emaka ja blastotsüsti arengu asünkroonsus tavaliselt vähendab blastotsüsti ellujäämust. On seejuures arvatud, et liiga vara ja liiga hilja emakasse saabuvad blastotsüstid on vähemedukamad implantatsiooniprotsessis. Hiirtel ja kariloomadel on näidatud, et isased blastotsüstid arenevad kiiremini kui emased. Seega, kui emakas on sünkroonis varasaabuvate blastotsüstidega, siis peaks sugude suhe kalduma isaste suunas, vastasel korral vastupidi.
c) Sooti erinev looteline suremus
Arvatakse, et isaste ja emaste looteline suremus on sőltuvalt hormoonide tasemest ning emaka toiduga varustamisest erinev. Vastavaid uurimusi on tehtud näiteks hiirtel ja sigadel ning näidatud, et isas- ja emasloodete suremused erinevad.
Kőigi nimetatud mehhanismide kohta on siiski tőendeid napilt ning need on pigem kaudsed. Lisaks sellele ei ole teada, kas sellisel viisil tekkinud sugude suhted on füsioloogiliste protsesside kaasprodukt vői emase vőime reguleerida oma järglaskonna sugude suhet, et maksimiseerida oma kohasust. On siiski vähemalt üks viis, kuidas emasloom saaks käitumuslikult muuta sugude suhet eelpool toodud füsioloogiliste mehhanismide läbi. See saab toimuda viljastamise aega valides. Kuna erinevate hormoonide tase ovulatsiooniperioodi jooksul muutub, on emasel potentsiaalselt vőimalik manipuleerida sugude suhet. On ka teada, et mitmesugused keskkonnafaktorid mőjutavad hormoonide taset, mis vőivad omakorda mőjutada sugude suhet.
Veel, kui enamus eelpool käsitletud sugude suhte kujunemise mehhanisme olid vähemal vői enamal määral adaptiivsed, siis sugude suhe vőib kujuneda ka mitteadaptiivselt, näiteks juhul, kui ühe sugupoole suremus on suurem. Näiteks lindude puhul on oletatud, et poegade suremus vőib olla kőrgem kui tütarde suremus.
KASUTATUD KIRJANDUS
* Charnov, E.L. 1979. The genetical evolution of patterns of sexuality: Darwinian fitness. American Naturalist 113: 465-480.
* Charnov, E.L. 1982. The theory of sex allocation. Princeton University Press, Princeton.
* Frank, S.A. 1990. Sex allocation theory for birds and mammals. Annual Review of Ecology and Systematics 21: 13-55.
* Godfray, H.C.J. 1994. Parasitoids: behavioral and evolutionary ecology. Princeton University Press, Princeton.
* Godfray, H.C.J., Werren, J.H. 1996. Recent developments in sex ratio studies. Trends in Ecology and Evolution 11: 59-63.
* Griffiths, N., Godfray, H.C.J. 1988. Local mate competition, sex ratio and clutch size in bethylid wasps. Behavioural Ecology and Sociobiology 22: 211-217.
* Hamilton, W.D. 1967. Extraordinary sex ratios. Science 56: 477-488.
* Hardy, I.C.W. 1994. Sex ratio and mating structure in the parasitoid Hymenoptera. Oikos 69: 3-20.
* King, B.H. 1987. Offspring sex ratios in parasitoid wasps. Quarterly Review of Biology 62: 367-396.
* Krackow, S. 1995. Potential mechanisms for sex ratio adjustment in mammals and birds. Biological Reviews of the Cambridge Philosophical Society 70: 225-241.
* Lummaa, V., Merilä, J., Kause, A. 1998. Adaptive sex ratio variation in pre-industrial human (Homo sapiens) populations? Proceedings of the Royal Society of London B 265: 563-568.
* Trivers, R.L., Willard, D.E. 1973. Natural selection of parental ability to vary the sex ratio of offspring. Science 179: 90-92.
* Waage, J.K. 1982. Sib-mating and sex ratio strategies in scelionid wasps. Ecological Entomology 7: 103-112.
* Werren, J.H. 1983. Sex ratio evolution under local mate competition in a parasitic wasp. Evolution 37: 116-124.
TEEMAKOHAST INTERNETIST
Frank, S.A. Sex allocation theory for birds and mammals (PDF-formaadis)
Frank, S.A. Sex allocation in solitary bees and wasps (PDF-formaadis)
Guynn, Jr.,D.C., Hamilton, R.J. The effects of adult sex ratio on reproduction in white-tailed deer (PDF-formaadis)
Johnson, D.H., Sargeant, A.B. Impact of red fox predation on the sex ratio of prairie mallards (HTML-formaadis)
KORDAMISKÜSIMUSED
1. Populatsioonis on mingi tunnuse suhtes kahesuguseid isendeid, neist üht tüüpi isendid on haruldased ja teist tüüpi isendid tavalised. Too bioloogiliselt mőttekad näited olukordadest, kus sagedusest sőltuv valik soosib haruldast tüüpi ja kus tavalist tüüpi!
2. Kirjelda hüpoteetilist looma, kellel ESS sugude suhe on isaste enamusega.
3. Teatavasti on imetajatel XX-kromosoomigarnituuriga emased ning XY isased. Milliseid järeldusi vőiks teha sugude suhet määravate geneetiliste elementide paiknemise kohta, kui a) emaste-isaste suhe populatsioonis on vőrdne; b) populatsioon on isaste enamusega (vőta eelduseks, et teised faktorid sugude suhte määramisel rolli ei mängi).
4. Seleta Fisher'i teooria loogikat sagedusest sőltuva valiku alusel.
5. Kuidas uuriksid Hamiltoni lokaalse paaritumiskonkurentsi hüpoteesi paikapidavust?
|