Toomas Tammaru loengukonspekt, kirjutatud jaanuaris 2000
Evolutsiooni vahetu jälgimine ja kirjeldamine geneetika terminites (dünaamiline lähenemine sissejuhatava loengu mõistes) on kahtlemata kõige vahetum viis evolutsiooni uurimiseks. Sellel on aga ilmseid puudusi:
- selline lähenemine on kasutatav vaid lühikese põlvkonnakestusega organismide puhul;
- selline lähenemine on kasutatav vaid tehistingimustes ja vaid harva looduses: vaid sellistel juhtudel, kus isendite järglased ka tõesti kinnipüütavad ja seeläbi uuritavad on.
Peale ilmsete puuduste on dünaamilisel lähenemisel ka vähem ilmseid puudusi. Nimelt pole lühiajaliste (nt. valiku-) katsete tulemused probleemitult ülekantavad pika aja jooksul toimunud evolutsioonile. Seda sellepärast, et populatsioonigeneetilise analüüsi tulemusel saadavad päritavuskoefitsientide ja geneetiliste korrelatsioonide väärtused iseloomustavad ikka vaid populatsioonide geneetilist struktuuri antud hetkel, see struktuur võib muutuda ja muutubki populatsiooni evolutsioneerumise käigus. Lühiajaliste valikukatsete tulemused pole seega ekstrapoleeritavad pikematele ajavahemikele.
Evolutsiooni protsessi asemel võime uurida evolutsiooni tulemust (staatiline lähenemine sissejuhatava loengu mõistes) ja lisatööd tehes teha (rohkem või vähem hästi) põhjendatud järeldusi protsessi kohta, mis antud tulemuseni on viinud. Evolutsiooniprotsessi rekonstrueerimine jääb muidugi alati suuremal või vähemal määral hüpoteetiliseks. Küll aga on põhimõtteliselt võimalik anda üsna selge vastus küsimusele, millised jõud hoiavad (evolutsioonilises mõttes) paigal vaadeldavat olekut, näiteks uuritava tunnuse vaadeldavat väärtust (stabiliseeriv valik).
Kirjeldatud lähenemine põhineb optimaalsusparadigmal (paradigma - keskse tähtsusega lähenemisviis miskis teadusharus, täpsemalt vt EE 7. kd.) ehk eeldusel, et uuritava tunnuse (või üldisemalt reaktsiooninormi) väärtused on evolutsiooni käigus jõudnud oma optimaalväärtusteni ehk siis väärtusteni, mis tagavad nende omanikule maksimaalse kohasuse (mõistagi on liiast eeldada, et populatsiooni kõigil isenditel on optimumväärtus, küll võib populatsiooni keskmine või täpsemalt modaalväärtus olla optimaalne).
Optimaalsusparadigma keskseks eelduseks oleva ülalmainitud väite kehtivusel on omakorda eeldusi. Esiteks, tuleb eeldada, et keskkond on püsinud pikka aega sedavõrd muutumatuna, et "tunnusel oleks olnud piisavalt aega oma optimumini jõuda" ehk siis optimum, mille poole on pikka aega evolutsioneerutud, on sama mis praegu. Eelduse kontrollimiseks (kasvõi mõtteliselt) on vaja kriitiliselt mõelda, kas valikusurved keskkonnas, milles uuritav tunnus kujunes on samad, mis toimivad looduses praegu. Ilmse näitena olukorrast, kus see nii ei ole olgu siinkohal toodud inimese elukäiguomaduste seletamine optimaalsusmudeli raames - väga paljusid meie omadusi ei saa pidada optimaalseiks (kohasust maksimeerivaiks) meie tänapäevases elukeskkonnas, küll võisid nad seda olla kiviajal.
Teiseks, tuleb eeldada, et geneetilise muutlikkuse puudumine pole takistanud optimumi saavutamist, ehk peaaegu sünonüümselt, evolutsiooni teel optimumini pole olnud piiranguid. Viimane eeldus on tavaliselt täidetud meetriliste tunnuste puhul, kui uuritav väärtuste vahemik jääb kaugele füüsikalistest piirangutest. Kindlasti on see eeldus täidetud, kui lisaks on uuritaval tunnusel praegugi kõrge päritavuskoefitsient (palju geneetilist muutlikkust). Teisiti võivad lood olla näiteks keeruliste morfoloogiliste tunnuste puhul. Nii pole ilmselt mõttekas väita, et meil pole kolmandat silma kuklas sellepärast, et sellise olemasolu kahandaks kohasust (poleks optimaalne). Ülimalt tõenäoliselt pole sellise fenotüübiga mutante kunagi esinenud (pole olnud vastavat geneetilist muutlikkust) ja looduslik valik pole nende kohasuse kohta oma sõna saanud öelda.
Kui on piisavalt põhjust arvata, et ülalloetletud eeldused on täidetud, võib hakata optimaalsuseeldust rakendades evolutsiooni uurima. Esimese sammuna püüame saada kvalitatiivset pilti faktoritest, mis uuritava tunnuse (reaktsiooninormi) eeldatavat optimumi paigas hoiavad. See tähendab, et püüame näidata, et populatsiooni keskmisest (või täpsemalt moodist) - mis on vastavalt eeldustele optimumiks - hälbiva tunnuse väärtusega isenditel on madalam kohasus kui keskmisele lähedasi väärtusi omavatel. Edasi püüame mõista, miks on madalam kohasus.
Lihtsaim viis selleks on vaatlus. Me võime leida loodusest hälbiva tunnuse väärtusega isendeid ja hinnata nende kohasust. Ehk õnnestubki meil näidata, et selliste isendite kohasus on väiksem. Siiski, sellise tulemuse interpreteerimine on problemaatiline. Seda seetõttu, et tunnuse hälbivate väärtustega loodusliku populatsiooni isendid erinevad keskmisest (moodist) tõenäoliselt ka muude omaduste poolest. Vaatlus ei võimalda väita, et hälbiva väärtusega isendeil oli madalam kohasus, sest neil oli uuritava tunnuse keskmisest erinev väärtus ehk teisisõnu, vaatlus ei võimalda üldjuhul tuvastada põhjuslikkust. Erijuht on selline, kus miskis lihtsas süsteemis saab põhjuslikkuse muud suunad miskil moel välistada.
Põhjusliku seose kindlakstegemise klassikaline meetod on katse. Katse sooritamise puhul ei kasuta me looduslikku varieeruvust, vaid me tekitame seda varieeruvust ise ehk me manipuleerime tunnuse väärtusi. Nii võime tagada, et uuritava tunnuse erinevate väärtustega rühmad ei erine muude tunnuste poolest. Evolutsioonilises ökoloogias huvipakkuvatest tunnustest on mitmed hästi manipuleeritavad. Klassikaline näide on siin lindude kurna suurus - munade lisamine pesadesse ja nende eemaldamine sealt on lihtne läbi viia (siinmail P. Hõrak ja I. Ots). Efektseks näiteks on va rahvuslinnu suitsupääsukese saba pikkus, mida eksperimentaator võib nii suurendada kui vähendada. Pääsukese saba morfoloogia manipuleerimisega on edukalt tegeletud Eestiski (M. Kose). Seevastu näiteks muna suuruse manipuleerimine on väga raske ülesanne.
Peale uuritava tunnuse väärtuste manipuleerimise on mõnel juhul võimalik manipuleerida ka selektiivset keskkonda. Nii on näiteks võimalik näidata, et piikide olemasolu kehal suurendab vesikirpude kohasust kiskjate olemasolul, seevastu vähendab seda keskkonnas, kus kiskjad puuduvad. "Kahepoolne" (tunnus ja keskkond) manipuleerimine olekski rangeimaks tõestuseks optimumi olemasolule ja faktorite olemusele, mis seda optimumi paigas hoiavad. Paraku tuleb praktikas sageli vähemaga leppida.
Peale optimumi paigas hoidva stabiliseeriva valiku kvalitatiivsele tõestamisele on ühe või teise valikufaktori tugevust võimalik ka kvantitatiivselt hinnata (mõõta). Kui optimumi mõjutavaid tegureid on palju ja eriti siis, kui nende mõju pole aditiivne (st olulised on koosmõjud - ühe faktori mõju sõltub teise mõjust), on tervikliku pildi saamiseks vaja matemaatilist mudelit. Sellise mudeli koostamiseks peame kõigepealt valima sobiva kohasuse praktilise definitsiooni, st mõõdetava suuruse, mille usume kohasusega piisavalt hästi korreleeruvat (nt järglaste arv, elumus - vt. sissejuhatav loeng). Seejärel peame matemaatiliselt väljendama oma arusaamise sellest, kuidas valitud kohasuse mõõt sõltub uuritava tunnuse väärtusest. Saadud mudelit matemaatiliselt analüüsides leiame, milline uuritava tunnuse väärtus maksimeerib kohasuse. Kui sellisel viisil leitud tunnuse optimumväärtus on lähedane looduses reaalselt vaadeldavale, annab see rõõmustava vihje sellest, et oleme koostanud õige mudeli ehk siis tunnuse evolutsioonist õigesti aru saanud. Siiski - sama tulemuse andvaid erinevaid mudeleid võib konstrueerida lõpmatu hulga, mudeli ja looduses vaadeldava kooskõla pole seega veel piisav tõestus asjast õigesti arusaamisele. Veenvaim tõestus on mudeli ennustusvõime. Asja tuum järgmine - mudel on esialgselt koostatud miskite kindlate keskkonnatingimuste jaoks ja seletab seda, mida teadsime enne mudelit koostama hakkamist. Selles avaldub mudeli kirjeldav võime. Mudeli ennustusvõime tähendab, et ta suudab ennustada, mida enne ei teadnud. Nii võime sobiva mudeli abil ennustada miski tunnuse väärtust miskis teises keskkonnas elavas populatsioonis, kus me selle tunnuse väärtusi mõõtnud pole. Kui asja looduses järgi uurides selgub, et mudel ennustas õigesti, on see juba parasjagu kaalukaks argumendiks mudeli (arusaamise) õigsuse poolt.
Ülalkirjeldatud viisil võime aru saada jõududest, mis tunnuse väärtusi paigal hoiavad. Sellisest teadmisest on kindlasti kasu mõistmaks, kuidas tunnus on sellise väärtuseni evolutsioneerunud. Selline tagasivaade eeldab aga paraku seda, et me teame, milline oli algolek (selline teadmine täieneb sedamööda, kuidas evolutsioonipuid rekonstrueeritakse) ja seda, et teame millised olid keskkonna olulised parameetrid siis ammu-ammu.
Näide 1. (siinkirjutaja oma praktikast). On kena hulk liblikaid, mille röövikud suudavad edukalt kasvada ja areneda vaid oma toidutaimede (lehtpuude) noortel lehtedel. Suur osa neist liblikaist lendab valmikuna (hilis)sügisel, kuigi kindlasti jõuaks areneda valmikuks juba juuliks (pole piirangut). Miks siis? Vaatluse all hallavaksik. Usun, et asja saab seletada optimaalsusmudeliga, sest 1) aastaaegadest tulenev selektiivne keskkond on ilmsestigi olnud pikka aega samasugune, suuri hiljutisi muutusi pole põhjust oletada (küll tuleks aga asja uurida võimalikult looduslikus keskkonnas, mitte linnaaias!); 2) tean, et hallavaksiku nukukestuses (nukuperioodi pikkuses) on palju geneetilist varieeruvust ja selle tunnuse võimet vastata valikule tõendab ka nukuperioodi erinev pikkus eri laiuskraadidel. Otsustasin katseliselt uurida, millised valikutegurid on vastutavad selle eest, et hallavaksik lendab septembri lõpus ja oktoobri alguses ja mitte suvel nagu enamik teisi liblikaid. Näen kolme peamist võimalust: a) sügise madalatel temperatuuridel suudab emane muneda rohkem kui kuumal suvel, mil eluiga on lühem; b) nukkude suremus on madalam kui munade suremus (üle jääv aeg tasub veeta pigem nuku kui munana!); c) valmikute elumus on sügisel kõrgem (kuna kiskjate arvukus on siis madalam). Vastavate katsete (valikusurvete katseline mõõtmine) tulemused veenavad, et võimalik põhjus b pole oluline (ei tekita märkimisväärset valikusurvet); põhjusel a on küll teatav mõju, kuid see ei küüni oma tugevuselt c tasemeni. Usun seega, et põhjus on eelkõige valmikute madalamas suremuses sügisel. Kirjeldatud võrdluses oli vaja kvantitatiivselt hinnata eri valikufaktoreid ehk siis täpsemalt nende mõju kohasusele. Selleks oli vaja miskist kohasuse praktilist definitsiooni ehk mõõtu. Seekord oli asi sellega suhteliselt lihtne - on põhjust arvata, et munetud munade arv (realiseerunud viljakus) iseloomustab emase hallavaksiku kohasust piisavalt täielikult.
Näide 2. (jälle siinkirjutaja oma praktikast). Olen püüdnud (optimaalsusparadigma raames) seletada sellesama hallavaksiku emase kehasuuruse determinatsiooni reaktsiooninormi (mõtlen funktsiooni, mis näitab, millise nukukaalu hallavaksik saavutab miski toidu kvaliteedi puhul). Kohasus (realiseerunud viljakus selle mõõduks) sõltub positiivselt nukukaalust ja negatiivselt arenguperioodi pikkusest (suhteliselt kaitsetu röövik on kauem kiskjatele eksponeeritud). Valmiku kohasuse sõltuvust nukukaalust on suhteliselt lihtne katseliselt määrata, seevastu röövikute suremus (kiskluse intensiivsus) on väga raskesti mõõdetav suurus. Seetõttu tuli teha suremuse kohta miskeid oletusi ehk siis analüüsida mudelit suremuse eri väärtuste korral (varieerides neid mõistlikkuse piirides) - seda räägin siin seepärast, et see on levinud võte modelleerimises - kui miskit parameetrit täpselt määrata ei õnnestu, tuleb uurida mudeli käitumist selle parameetri hüpoteetiliste realistlike väärtuste korral, kui kvalitatiivsed järeldused selle muutuja väärtustest ei sõltu, polegi selle muutuja tegelikest väärtusest ja nende mitteteadmisest suurt lugu. Vastava mudeli analüüs näitas, et hallavaksiku emase kehasuurust ei saa pidada optimaalseks - kui röövik kasvaks kauem, suureneks isendi kohasus. Asjal kaks võimalikku seletust - 1) minu mudelist on midagi olulist puudu; 2) antud tunnus pole tõepoolest ökoloogilises kontekstis optimaalne, asjasse on segatud mingi piirang, mis ökoloogiliselt optimaalset väärtust saavutada ei lase. Minule meeldib uskuda teist varianti.
Näide 3. (välja mõeldud näide matemaatilisest mudelist, päris mudelid on siinkohal esitamiseks enamasti liiga keerulised).
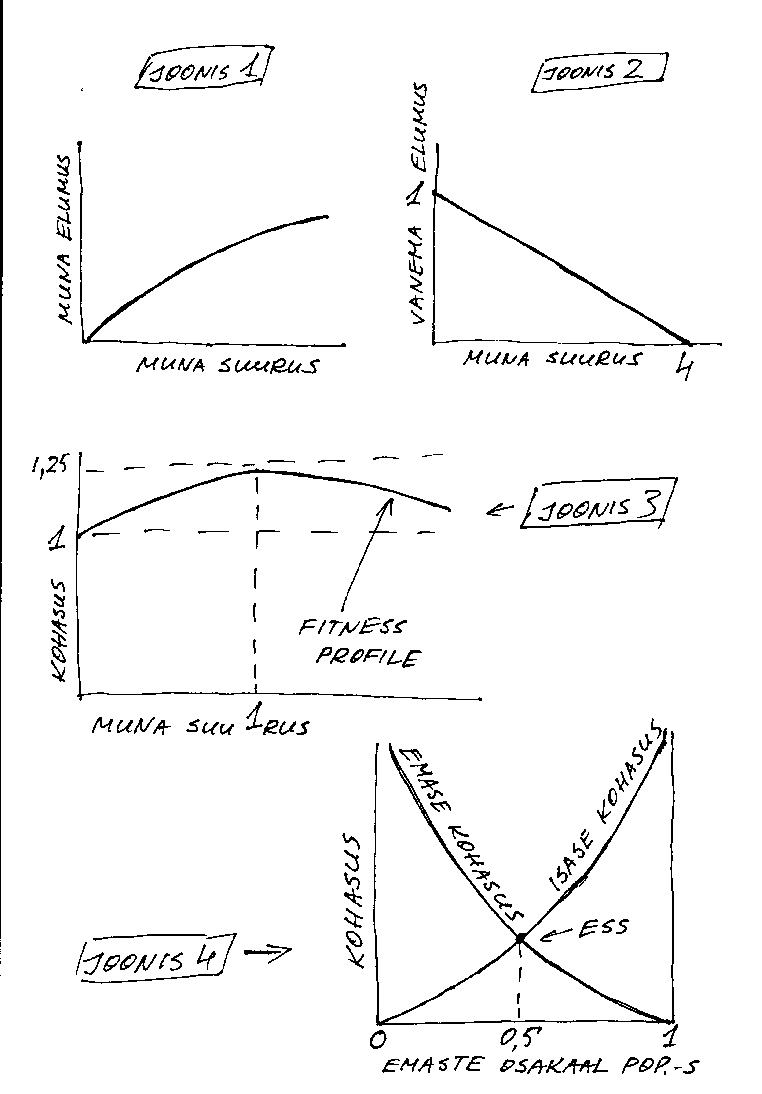
Olgu uuritavaks tunnuseks miski ussikese muna suurus, ülesandeks on hinnata suuruse optimumi. Mängime, et ussikese muna elumuse lm sõltuvust muna suurusest x iseloomustab (katseliselt kindlaks tehtud) kõver (joonis 1), mis on matemaatiliselt (ligikaudselt, muidugi) kirjeldatav funktsiooniga
lm=1 - 1/(x+1)
ja mängime edasi, et muna munenud vanemussikese elumus lv järgmise paljunemiskorraga sõltub munetud muna suurusest järgmiselt (joonis 2), mis on kirjeldatav sirge võrrandiga
lv=1-(x/4)
Kui nüüd lisaks oletada, et kohasus ei sõltu millestki muust, võime muna munenud ussikese kohasuse esitada selle kahe komponendi summana,
w = lm+ lv = 2 - 1/(x+1) - x/4
Nagu funktsiooni maksimumi ikka leitakse, leiame funktsiooni tuletise ja võrdsustame selle nulliga.
dw/dx = 1/(x+1)2 - 0.25 = 0
Lahendades saadud võrrandi leiame, et kohasus on maksimaalne muna suuruse x=1 korral (joonis 3). Antud näites lahendasime ülesande analüütiliselt, st funktsioonianalüüsi meetodeid kasutades. Konkreetsete juhtude uurimisel on empiiriliselt saadud funktsioonid enamasti sedavõrd keerukad, et ülesande analüütiline lahendamine ei ole võimalik või vähemalt mitte otsatarbekas. Sellistel juhtudel tuleb tunnuse optimaalväärtus (või muu meid huvitav parameeter) leida numbriliselt ehk siis lihtsalt öeldes - proovimise teel, kõigi mõistlike parameetrikombinatsioonide süstemaatilise läbiproovimise teel. Proovimisega tegeleb muidugi arvuti. Teoreetikuid huvitab aga analüüsida ja tõestada asju üldjuhul (et näiteks kui see kõver on selliste omadustega ja too teistega, kas siis optimaalne reaktsiooninorm on sellise või teise kujuga), sellisel puhul on analüütiline lähenemine muidugi eelistatud.
Näidete juurest tagasi üldisemale tasemele. Kõik eelpool toodud näited on liigitatavad nn. lihtsa optimeerimise alla (minu väljend, mitte üldtunnustatud termin). Lihtsa selles mõttes ja sellepärast, et selektiivne keskkond (tunnuse väärtustele valikusurvet avaldavate
tegurite kogum) ei sõltu tunnuse väärtusest populatsioonis. Nii ei sõltu ka tunnuse optimaalväärtus sellest, milline on selle tunnuse väärtuste jaotus populatsioonis. Näiteks pole ehk põhjust arvata, et munakoore paksuse optimum sõltuks sellest, milline on munakoore
paksuse väärtuste sagedusjaotus populatsioonis. Alati see nii aga ei ole. On olukordi, kus tunnuse evolutsioon populatsioonis (tunnuse väärtuste sagedusjaotuse muutus) mõjutab selektiivset keskkonda, muudab seega ka väärtuse optimumi. Selliste olukordade kohta leiab ehk kõige hõlpsamini näiteid käitumisökoloogia vallas. Nii sõltub miski käitumisviisi adaptiivsus enamasti sellest, mismoodi käitub käitumisakti partner. Nii on agressiivne käitumine kahtlemata eelistatud järeleandliku ees, kui agressiivsus on populatsioonis haruldane. Eeldades, et käitumisviisid on geneetiliselt determineeritud, hakkab loodusliku valiku tõttu sellises populatsioonis tõusma agressiivselt käitujate osatähtsus. Kahe agressiivselt käituja kokkupõrkel võivad aga olla kurvad tagajärjed mõlemale ja agressiivse käitumise piisava sageduse korral populatsioonis polegi enam kuigi adaptiivne sellist käitumisviisi omada (käitumise evolutsioon populatsioonis põhjustas selektiivse keskkonna muutumise). Agressiivse käitumise miski sageduse juures on tasakaalupunkt, tasakaalupunktis on võrdselt adaptiivne käituda järeleandlikult või agressiivselt.
Selliste olukordade analüüsiks ei saa enam kasutada lihtsa optimeerimise loogikat, vaja on ESS-loogikat. ESS on lühend väljendist "evolutsiooniliselt stabiilne strateegia". Sõna "strateegia" sisaldub selles väljendis ehk just seetõttu, et sellist lähenemist rakendati kõigepealt just käitumisega seotud optimaalsusülesannete lahendamisel. Selle lähenemise rakenduspiirkond on märksa laiem ja alati ei sobi uuritavat tunnust strateegiaks nimetada, seega üldisem oleks sehk rääkida evolutsiooniliselt stabiilsest seisundist, no ESS igal juhul.
Süsteem, mis on evolutsiooniliselt stabiilses olekus, ei evolutsioneeru (loodusliku valiku läbi). Süsteem, mis ei ole ESSs evolutsioneerub ja saavutab üldiselt varem või hiljem ESSi (siiski on teoreetilisi mudeleid, mis ennustavad, et mõnikord võib ESS olla teatud lähteolekust saavutamatu). Olukorrale, kus ESS olukorras on populatsioonis esindatud mitmed "strateegiad" (mixed ESS) on omane, et kõigi strateegiate kohasus on võrdne (kui nii poleks, läheks ju evolutsioon lahti).
ESSi modelleerimine on oluliselt keerukam kui lihtsa optimeerimise põhimõttele tuginevad mudelid. Tõestamaks, et miski olukord on ESS, kasutatakse sageli meetodit nimega invadability analysis. Asja tuum on selles, et uuritakse, kas miski uue "sissetungija" kohasus võib olla kõrgem populatsiooni isendite keskmisest kohasusest. Kui on, levib sissetungija populatsioonis, milline olukord tõestab, et tegu polnud ESSga. See sissetungija võib olla alleel, genotüüp või fenotüüp; sõltuval sellest, kas küsimus on püstitatud ja mudel koostatud alleelide, genotüüpide või fenotüüpide tasemel. Kui õnnestub tõestada, et miski sissetungija ei suuda populatsioonis levida, on tegu ESSga. Vastavalt sellele, kui laia genotüüpide hulga kohta väide kehtib, võib eristada kolmel tasemel ESSe:
- sisemine stabiilsus (internal stability). Populatsioon on stabiilne populatsioonis olemasolevate genotüüpide sageduse muutuse suhtes, ehk siis mis iganes ja mis põhjusel ka ei
juhtuks populatsioonis olemasolevate genotüüpide sagedusega, taastub endine st ESS olek (uute mutatsioonide kohta ei väideta midagi);
- piiratud välisstabiilsus (limited external stability). Süsteem on stabiilne mitte ainult populatsioonis olemasolevate genotüüpide sageduse muutuste suhtes, vaid on resistentne ka teatud (st. defineeritud) hulga mutantide sissetungi suhtes. Ehk me siis küsime, kas nii- ja naasugused mutandid, mida me parasjagu ette kujutada tahame või oskame, suudavad populatsioonis levida - kui nad seal tekkima peaksid. Eitav vastus tähendab, et populatsioon on ESS olukorras kõnealuse stabiilustaseme mõttes;
- välisstabiilsus (external stability). Süsteem on resistentne mistahes mutandi sissetungi suhtes. Mõistagi mõtleme siin mutatsioone, mis mõjutavaid neid tunnuseid, mille evolutsioonilist stabiilsust parasjagu uurime.
Kui uurime lühiajalisi evolutsioonilisi protsesse, võime opereerida sisemise stabiilsuse
mõistega. Ehk meile piisab, kui uurime, kas miski olemasolev alleel või genotüüp võib populatsioonis levida ja olemasolevat tasakaalu paigast nihutada, uute mutatsioonide tekkimise võimalusega võime mitte arvestada. Mida pikemaaegsem evolutsioon meid huvitab, seda rohkem nihkub meie huvi välissstabiilsuse suunas.
Käitumisega seotud ESSide temaatikat ja näiteid käsitletakse käitumisökoloogia loengus. ESS-tüüpi lähenemine on kasutatav peale populatsioonisiseste käitumis- ja muude strateegiate probleemide näiteks eri liikide koevolutsiooni uurimisel. Antagonistlike suhete puhul võib kergesti tekkida olukord, kus ESSi polegi ja süsteem on pidevas evolutsioonilises liikumises. Nii on parasiidil pidevalt kasulik peremeest üllatada näiteks uue biokeemilise nüansiga, mis teeb peremehel parasiidi vastu võitlemise raskemaks, peremehel on seejärel kasulik uus nüanss "üle kavaldada". Tegemist on pideva "evolutsioonilise võidujooksuga" (nn. Red Queen-fenomen). Peremees-parasiit võidujooksus edukana püsimiseks on ilmselt kasu sugulisusest, kuna sugulisus genereerib pidevalt uusi ja vastaspoolele üllatavaid genotüüpe. Sugulisuse laia levikut looduses selle värgi kulukusele vaatamata on viimasel ajal hakatud põhjendama just peremees-parasiit võidujooksuga.
Lõpuks üks ehk mittevähese tähtsusega kvantitatiivne näide ESS olukorrast. Nimelt näitame, et sugupoolte suhe 1:1 on üldjuhul ESS. Veidi järgi mõeldes võib igaüks veenduda, et rangelt biparentaalsete (igal isendil on kaks geneetilist vanemat) lahksooliste elukate populatsioonis on kõigi isaste summaarne kohasus võrdne kõigi emaste summaarse kohasusega. Seega on ühe isase keskmine kohasus võrdeline isaste osatähtsuse pöördarvuga populatsioonis. Seega, kui populatsioonis on emaste ülekaal, on kasulikum (st tagab kõrgema
kohasuse) olla isane ja vastupidi. Kuna aga soo muutumine täiskasvanud loomal pole enamasti võimalik (erandeid on), siis on ehk olulisem tähelepanek, et populatsioonis, milles on emaste enamus, on kasulik toota poegade ülekaaluga järglaskondi. Eeldusel, et sugupoolte suhe järglaskonnas on (vähemalt osaliselt) geneetiliselt determineeritud, hakkab emaste ülekaaluga populatsioon evolutsioneeruma emaste ülekaalu vähenemise suunas. See protsess peatub, kui isaste ja emaste osakaal populatsioonis on võrdne, sellises olukorras on emase ja isase oodatavad kohasused võrdsed: tegu on ESSiga (joonis 4).
Küsimusi kordamiseks ja eksamiks
1. Optimaalsusparadigma, selle rakendamise eeldused.
2. Too näide miski looma miskist tunnusest (soovitavalt mitte ülalesitatud näidete hulgast), mille väärtuse usud olevat optimaalse. Kuidas uuriksid, millised tegurid põhjustavad stabiliseerivat valikut? Kas tunnuse väärtust ja/ või selektiivset keskkonda on võimalik manipuleerida? Milline oleks siin manipulatsiooni eelis vaatlusega võrreldes?
3. Mõtle välja näide probleemist evolutsioonilise ökoloogia vallas, kus läheks vaja matemaatilist modelleerimist (mudelit pole siiski vaja koostada).
4. Too näide tunnusest, mille väärtuse evolutsioon muudab tunnuse optimumi. Milline võiks olla ESS?
5. Populatsioonis on 60% isenditest tunnuse A väärtus a1 ja 40% isenditest väärtus a2 (teame, et väärtus on geneetiliselt määratud). Mil viisil on võimalik uurida, kas vaadeldav seisund on ESS?
7. Seleta mõisteid: sisestabiilsus, piiratud välisstabiilsus, välisstabiilsus.
6. Seleta, miks evolutsioneerub emaste ülekaaluga populatsioon emaste ülekaalu vähenemise suunas.
{kind=link}